
ABSTRACT
A transfer-prepared specimen of Lepidotes (Actinopterygii, Lepisosteiformes) from the Posidonia Shale Formation of south Germany is tentatively identified specifically as L. gigas. The genus and its species are insufficiently defined and in need of revision. The new specimen reveals details of the cranial and pectoral girdle skeletal architecture and previously unknown osteological features of Lepidotes, such as a large foramen on the maxilla and the formation of two tooth-bearing clavicle elements (serrated appendages). Maxillary foramina are also known in the related contemporaneous lepisosteiform Mengius and possibly served for passage of nerve branches supplying either a maxillary pit-line or cutaneous sense organs (taste buds) in a thickened upper lip or barbels. The clavicle elements agree morphologically with those of the Recent Amia. Their function remains obscure.
INTRODUCTION
Agassiz (Citation1832) described under the name of Lepidotes gigas large fusiform fishes from the Lower Jurassic “Lias Formation” (Posidonia Shale Formation) in southern Germany of which he found beautiful specimens in the private Hartmann collection. E. G. F. von Hartmann (1767–1851) was a physician in the city of Göppingen 40 km east of Stuttgart, southern Germany, and then a well-known fossil collector. According to Agassiz (Citation1832) the specimens came from the locality Ohmden, a village near Holzmaden in the foreland of the Swabian Alb. A little later, specimens of this fossil fish were also studied by F. A. Quenstedt (Citation1847) from the University of Tübingen who thought them to be conspecific with Lepidotes elvensis (Blainville, Citation1818) from the Lower Jurassic of France. More recently, López-Arbarello (Citation2012) showed that L. gigas Agassiz, Citation1832 and L. elvensis (Blainville, Citation1818) represent different species disagreeing in cranial shape and architecture and in the morphology of some cranial ossifications.
Even though species of Lepidotes and closely related genera (see López-Arbarello, Citation2012; Thies et al., Citationin press) belong to the most prominent fossil fishes with specimens on exhibition in many museums around the world the type species L. gigas is still insufficiently known and defined. Since Agassiz (Citation1832, Citation1833, Citation1834, Citation1837), Quenstedt (Citation1847, Citation1852, Citation1858), and Woodward (Citation1895) only little was added to the knowledge of L. gigas. López-Arbarello (Citation2012) proposed an amended diagnosis of the genus Lepidotes Agassiz, Citation1832 and specified some morphological and anatomical differences between L. gigas and L. elvensis. However, despite an immense amount of material housed in public collections a comprehensive description of L. gigas following the rules of the International Code of Zoological Nomenclature (Citation1999) is still missing. In this paper previously unknown cranial and pectoral girdle skeletal features of L. gigas are described.
MATERIALS AND METHODS
Material—The material consists of one nearly complete, articulated specimen of the fossil fish Lepidotes gigas Agassiz, Citation1832 missing the caudal fin. The fish was preserved in a calcareous nodule, which was split during recovery into halves to produce part and counterpart. The nodule was split in a way that part and counterpart enclosed the left and right half of the fish corpus, respectively. Both parts were subsequently prepared separately. The part of the specimen was previously studied and published with regard to its preserved stomach contents (Thies et al., 2019).
Preparation—Details of preparation were already given in Thies et al., (Citation2019) and will only be briefly summarized here. The specimen (part) was transfer-prepared according to Rixon (Citation1976) and Rutzky et al. (Citation1995). After preparation it shows the left side. Slight dislocation of cephalic and pectoral girdle ossifications suggests that this side was turned towards the water body during taphonomic processes while the right side rested on the sediment.
Osteological Description—In the morphological description of the cephalic skeleton the nomenclature for cranial ossifications proposed by Schultze (Citation2008) is applied, which at present is the best expression of homology between individual bones of actinopterygians and sarcopterygians within Osteichthyes. The terminology for the description of the sensory line system follows Devillers (Citation1958) and Thies (Citation1989a, Citationb, Citation1991).
Institutional Abbreviations—FWD, Fossilienmuseum im Werkforum Dotternhausen, Holcim (Süddeutschland) GmbH, Dotternhausen, Germany.
SYSTEMATIC PALEONTOLOGY
Subclass ACTINOPTERYGII Cope, Citation1887
Order LEPISOSTEIFORMES Hay, Citation1929
Family LEPIDOTIDAE Agassiz, Citation1833
Genus LEPIDOTES Agassiz, Citation1832
LEPIDOTES GIGAS Agassiz, Citation1832
( and )
FIGURE 1. Lepidotes gigas Agassiz, Citation1832, FWD_1098 (part). A, transfer-prepared specimen in left lateral view, dusted with ammonium chloride for photography; B, cephalic and pectoral girdle skeleton of the same; C, drawing of cephalic and pectoral girdle skeleton. Abbreviations: Ang, angular; Art, articular; Cl, cleithrum; Cla, anterior clavicle element; Clp, posterior clavicle element; Dpal, dermopalatine; Dspl, dentalosplenial; Epi, epiotic; Ex, extrascapular; Exo, exoccipital; hpl, horizontal pit-line of the cheek; Hym, hyomandibula; Io, infraorbital; Iop, interoperculum; mf, maxillary foramen; Mtp, metapterygoid; Mx, maxilla; Na, nasal; Op, operculum; Pa, parietal; p.bas, processus basipterygoideus; pccll, pores of the canal for the cephalic lateral line; pcio, pores of the canal for the infraorbital sensory line; Pcl, postcleithrum; pcoio, pores of the canal for the otic section of the infraorbital sensory line; pcpocll, pores of the canal for the postotic section of the cephalic lateral line; pcsc, pores of the canal for the supratemporal commissure; pcsop, pores of the canal for the supraorbital sensory line; pmdc, pores of the mandibular canal; Pmx, premaxilla; poc, pores of the “oral” canal; Pop, preoperculum; Ppa, postparietal; ppc, pores of the preopercular canal; ppl, posterior pit-line of the skull roof; Pro, prootic; Pt, posttemporal; Qu, quadrate; Quj, quadratojugal; Sang, surangular; Smx, supramaxilla; Sob, suborbital; Sop, suboperculum; Stta, supratemporotabular; Sym, symplectic; Vo, vomer; vpl, vertical pit-line of the cheek; (l), left; (r), right.
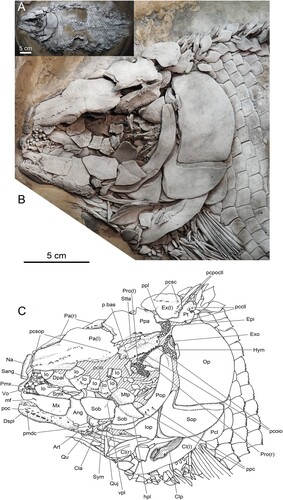
FIGURE 2. Lepidotes gigas Agassiz, Citation1832, FWD_1098 (part). A, skull and pectoral girdle in left lateral view, white squares indicate position of details shown in B, C, and D, dusted with ammonium chloride for photography; B, left maxilla showing a large slot-like foramen on its anterior part; C, anterior clavicle element (anterior serrated appendage) being intermingled with remains of branchiostegal rays; D, posterior clavicle element (posterior serrated appendage) lying on external surface of ventral arm of left cleithrum.

Referred Specimen—FWD_1098 (part).
Locality—Working quarry for cement at the village of Dotternhausen on the western slope of the Swabian Alb, south Germany. Grid reference: TK 25 Geislingen 7718, 48°13′50.16′′N, 8°46′21.21′′E.
Age—Early Jurassic, early Toarcian, lower falciferum zone, lower exaratum subzone.
Depository—Fossilienmuseum im Werkforum Dotternhausen, Holcim (Süddeutschland) GmbH.
Note—The specimen was previously identified as Lepidotes sp. (Thies et al., 2019). It is considered here as belonging to Lepidotes gigas Agassiz, Citation1832. However, this identification is only provisional because L. gigas was never diagnosed properly. The description and figures of L. gigas by Agassiz (Citation1834, Citation1837) have only little significance because he based the description on a mixture of specimens from southern Germany, France, and England. Furthermore, in his works of Citation1833 and Citation1837, Agassiz already synonymized L. gigas with L. elvensis (Blainville, Citation1818). This view was adopted later by Quenstedt (Citation1847) who gave the name L. elvensis priority over L. gigas. The whereabouts of the originals of Agassiz’ figures (Citation1834:pls. 28, 29) are unknown. In contrast, a large part of the material Quenstedt (Citation1847) based his study of Lepidotes on still exists in the Tübingen University paleontological collection. This material is of the same age and comes from the same locality as Agassiz’ south German specimens. It obviously belongs to L. gigas and for the moment can serve as comparative material for the identification of new L. gigas specimens.
Specimen FWD_1098 agrees with the specimens of L. “elvensis” (= L. gigas) in Quenstedt (Citation1847:pl. 1, figs. 1, 2, and pl. 2, fig. 1) with regard to morphology and proportions of the skull roofing and opercular bones. It also agrees concerning the number and shape of suborbital elements (two of which have become detached during preparation in FWD_1098 (part), are kept separately and are therefore not shown in and ). Finally, all specimens have a tritoral dentition. Considering body shape, López-Arbarello (Citation2012) showed that L. gigas has a squat skull, which is more slender in L. elvensis. In this respect FWD_1098 also corresponds to L. gigas.
Further species of Lepidotes occurring in the Toarcian of Europe are L. semiserratus Agassiz, Citation1837 and L. buelowianus Jaekel, Citation1929. From the diagnosis of L. semiserratus in Woodward (Citation1897) clear differences from L. gigas are not evident. L. semiserratus may therefore be a younger synonym of L. gigas. Finally, L. buelowianus is a nomen dubium (Thies et al., Citationin press). To clarify the taxonomic questions related to L. gigas a revision and redescription of the species is in urgent need.
DESCRIPTION OF CRANIAL AND PECTORAL GIRDLE OSTEOLOGY
General Comments—An overview of the cranial and pectoral girdle osteology of the specimen is given in . At the end of the taphonomic processes the fish underwent after dying the body came to rest on the sediment with the left side up. This side was therefore exposed to bottom water currents and scavengers who attacked the carcass and fed on soft tissues of the head. In this way they disturbed and disarranged the skeleton of the skull and shoulder girdle and even removed individual bones. This refers in particular to the circumorbitals and the underlying ossifications. The two suborbitals posterior to the orbit are, however, not missing but are kept separately because they lost connection to the fossil during preparation.
Endochondral Neurocranium
The endochondral neurocranium has become disintegrated with some of its ossifications being partly visible in the specimen (). In the photograph in , the basisphenoid is also visible in the center of the orbit. In the schematic line drawing in it is omitted and replaced by hatching.
Dermal Neurocranium
Within the dermal neurocranium the skull roof is formed by a large parietal anteriorly, a postparietal in the middle, and a single extrascapular posteriorly (). A supratemporotabular completes the skull roof postero-laterally (). These bones bear different sections of the cephalic lateral line canals. Pit-lines can hardly be traced on the parietal, postparietal, and supratemporotabular, with the exception of the hind portion of the posterior pit-line of the skull roof, which extends a short way onto the extrascapular (). Rostrally the nasal is only partially preserved, and the rostral is not preserved at all in the specimen ().
The cheek is covered by the circumorbitals arranged around the orbit. The infraorbital ring () is incompletely preserved leaving the exact number of infraorbitals open. The supraorbitals and the dermosphenotic are missing. As well, the number of anterior infraorbitals remains unknown. There are four suborbitals, two of which are still present on the specimen (). The other two are kept separately. The infraorbitals bear the canal for the infraorbital sensory line (); the suborbitals below the orbit show grooves for the vertical and horizontal pit-line of the cheek ().
Two bones contributing to the formation of the palatal roof can be observed—the vomer anteriorly and a part of the parasphenoid posteriorly. The vomer is a paired ossification with the left vomer being shifted a little posteriorly after death (). The vomers bear a very strong tritoral dentition on their external surface. Of the parasphenoid only the (presumably right) basipterygoid process is preserved ().
Mandibular Arch and Associated Ossifications
The upper jaw comprises the premaxilla anteriorly and the maxilla and one supramaxilla posteriorly (). Premaxilla and maxilla bear teeth with those of the premaxilla being much stronger than those of the maxilla. A large, slot-like foramen opens on the anterior portion of the maxilla and supposedly served for the passage of nerves and/or vessels through the bone (). The hind border of the maxilla is damaged. Further ossifications associated with the upper jaw include the dermopalatine and the metapterygoid (). A single row of styliform teeth is present along the ventral edge of the dermopalatine. The metapterygoid is a concave lamina of bone with smooth surfaces. Of the quadrate only the condyle for the articulation of the articular bone of the lower jaw is visible (). Next to the quadrate condyle is the quadratojugal, which is free and dislocated a little posteriorly in the specimen ().
The lower jaw consists of a compound bone laterally, the dentalosplenial, to which the articular and the angular are sutured posteriorly (). The suture between the dentalosplenial and angular is developed only posteriorly and vanishes anteriorly. The surangular is shifted a little posteriorly (). Two canals are present on the lower jaw––the “oral” canal antero-dorsally and the mandibular canal ventrally, the pores of which extend along the ventral edge of the dentalosplenial and the articular (). The “oral” canal was previously considered to house an additional sensory line on the lower jaw (Thies, Citation1989a, Citationb, Citationc). Cavin and Suteethorn (Citation2006) found the “oral” canal to be present in different patterns of formation in a series of fossil ginglymodian taxa. Recently, Leuzinger et al. (Citation2020) confirmed that the “oral” canal is independent from the mandibular canal by μ-CT studies of an isolated lower jaw of Scheenstia sp. from the Upper Jurassic of Switzerland. They assumed that this canal rather served for the passage of rami of the trigeminal-facial nerve innervating (hypothetical) dermal sensory structures in thickened lips or barbels. There are no traces of the mandibular pit-line discernible on the lower jaw.
Hyoid Arch
Only the dorsal end of the hyomandibula is visible on the specimen (). The rest of the bone is concealed by the overlying preoperculum and the opercular apparatus. The opercular series is dislocated posteriad and its elements are slightly pulled apart from one another. It consists of a large, roughly rectangular operculum, a suboperculum with a prominent antero-dorsal process, and a triangular interoperculum (). In front of the opercular series lies a curved preoperculum being traversed by the preopercular sensory canal (). The symplectic, which is still in touch with the quadrate condyle, connects the hyoid arch with the mandibular arch (). Some branchiostegal rays are preserved but are not labelled in .
Pectoral Girdle
The posttemporal is of triangular shape and is rotated anticlockwise for nearly 90° so that the recess at its ventral border for the articulation with the dorsal process of the supracleithrum opens posteriad (). The canal for the cephalic portion of the lateral line passes through the posttemporal (). The supracleithrum and the dorsal branches of the cleithra are covered by the operculum and suboperculum, respectively, and are not observable.
The specimen displays the ventro-rostral arms of both cleithra, which are wide and convex and end in tips. The left cleithrum shows the lateral face of this arm (). A thin, strip-like lamina of bone bearing numerous longitudinal rows of tiny denticles and being only loosely attached to the lateral face of the left cleithrum represents the posterior clavicle element (posterior serrated appendage) (). A second but smaller such bony lamina among some disarranged branchiostegal rays (not labelled) is the anterior clavicle element (anterior serrated appendage) (). The rostro-ventral arm of the right cleithrum becomes visible a little dorsally and rostrally of the left cleithrum (). It displays its medial surface which carries two pronounced ridges.
DISCUSSION
In the formation of the cranial and pectoral girdle skeleton and in skull shape the specimen studied here closely resembles the specimens of Lepidotes gigas from the lower Toarcian of south Germany described by Quenstedt (Citation1847) and López-Arbarello (Citation2012). They are therefore considered as conspecific. Two of the skeletal features present in specimen FWD_1098 require, however, a more detailed consideration.
Foramen on the Maxilla
The maxilla of the specimen is broken (, ). Anterior to the rupture line opens a large slit-like foramen in the middle of the lateral surface of the ossification having its longitudinal axis oriented horizontally (, ). Some smaller pores are present on the postero-ventral portion of the bone. Functional explanations of the openings, in particular of the large anterior one, include two alternatives:
Maxillary Pit-line—In size, shape, and distribution the maxillary foramina agree with similar pores on the lateral face of the maxilla of the fossil lepisosteiform Mengius acutidens gen. et sp. nov. from the lower Toarcian of northeast Germany (Thies, Citation1989b:695, 697, figs. 2, 3; Thies et al., Citationin press). In M. acutidens these pores were interpreted as indicating the presence of a maxillary sensory line (pit-line) in the soft tissue covering the maxilla. The pores would function as passage for nerve and vascular supply of the sensory line. The lack of a characteristic groove on the surface of the maxilla was explained by a thickening of epidermal tissue on the maxilla caused by formation of a lip (for explanation of groove formation on the bone surface beneath pit-lines see Allis, Citation1889). The reasoning for the presence of a maxillary pit-line in M. acutidens includes (Thies, Citation1989b):
A maxillary sensory line can be present in fishes as demonstrated by representatives of the fossil genus Ophiopsiella Lane and Ebert, Citation2015. The sensory canal in the maxilla of Ophiopsiella was shown by Bartram (Citation1975) and Lane and Ebert (Citation2015) to join the infraorbital canal rostrally, and was therefore considered to be a branch of the infraorbital canal.
Among the sensory lines enclosed in canals and being present on the skull of fishes and amphibians, Stensiö (Citation1947) listed the supramaxillary line, which extends above and behind the maxilla and connects the preopercular with the infraorbital line. According to Stensiö (Citation1947) the supramaxillary line is developed in actinopterygians more superficially and is expressed as the horizontal pit-line of the cheek. A pit-line observed on the maxilla of some Paleozoic and early Mesozoic chondrosteans is believed to represent the anteriormost portion of the horizontal pit-line of the cheek (Gardiner, Citation1984; Lehman, Citation1966; Stensiö, Citation1947). The horizontal pit-line of the cheek is visible in L. gigas on the suborbital covering the ventral arm of the preoperculum (). In contrast to Bartram (Citation1975) a possible pit-line on the maxilla of L. gigas (and M. acutidens) could therefore also represent an anterior part of the horizontal pit-line and be homologous to the anterior part of the supramaxillary line.
Some taxa of the teleost families Esocidae, Cyprinidae, Siluridae, Gadidae, and Percidae possess superficial neuromasts forming an additional sensory line ventrally to the infraorbital line (Coombs et al., Citation1988; Jakubowski, Citation1966, Citation1967; Lekander, Citation1949; Pehrson, Citation1944). Different terms were used for this line: infraorbital pit-line (Pehrson, Citation1944), accessory infraorbital line (Lekander, Citation1949), ventral infraorbital line (Coombs et al., Citation1988), or simply infraorbital line (Arratia & Huaquín, Citation1995). The homology of this line is unclear. Coombs et al. (Citation1988) supposed a homology with the ventral infraorbital canal of holocephalans and actinistians.
Passage for Branches of the Trigeminal and/or Facial Nerves—Alternatively, the large foramen on the maxilla of L. gigas may be related to the course of nerves supplying the soft tissues of the upper jaw. In Amia the cheek and upper jaw is innervated mainly by rami of the trigeminal and facial nerves: ramus maxillaris superior trigemini, r. buccalis facialis, and r. mandibularis externus facialis (Allis, Citation1897). Of these, branches of the r. maxillaris superior trigemini are intimately connected with the maxilla and surrounding soft tissues. It seems therefore possible that one or more of the branches of the r. maxillaris superior trigemini passed from internally through the large anterior foramen of the maxilla to innervate soft tissue at or near the maxilla externally. Leuzinger et al. (Citation2020) considered the “oral” canal in the lower jaw of Scheenstia sp. as passage for branches of the trigeminal and facial nerves supplying sense organs embedded in thickened lower lips or barbels. Accordingly, Lepidotes might have possessed cutaneous sense organs such as mechano- or chemoreceptors in or on thickened upper lips or barbels, which were innervated by branches of the trigeminal and/or facial nerves. Among Recent teleosts a close relationship between the maxilla and barbels is for instance known in Ameiurus (Siluriformes) in which the maxilla supports the elongate, laterally extending maxillary barbel (Kindred, Citation1919). Cutaneous sensory organs, in particular taste buds, are embedded in large numbers in the skin of the lips and barbels of, for example, many siluriform species (Arratia & Huaquín, Citation1995). These external taste buds are innervated by branches of the facial nerve (Finger, Citation1988).
Clavicle Elements (Serrated Appendages)
There are two clavicle elements present in the pectoral girdle of FWD_1098––an anterior and a posterior one (, ). They do not appear to have been firmly attached to the pectoral girdle. The posterior element () lies on the lateral surface of the ventral arm of the cleithrum. The anterior element () is intermingled with some disarranged branchiostegal rays behind the articular of the lower jaw and seems to have been shifted rostrally during fossil diagenesis. Both elements agree in being built by thin laminae of dermal bone and in carrying tiny, hook-like denticles on their lateral (external) surfaces. The denticles are packed densely and cover nearly the entire surface in both elements. Further agreement can be observed in denticle morphology. All are multicuspid having three or four apices, respectively. The apices are arranged in a row so that the denticles are of comb-like appearance. Differences exist in size and shape of the clavicle elements and in the orientation of the denticles. The posterior clavicle is larger, narrow and strip-like with pointed dorsal and ventral tips. It reaches at least 2.3 cm in maximum longitudinal diameter. The anterior element is smaller and oval shaped. It is less than half in longitudinal diameter compared with the posterior element. Denticles form numerous rows on both elements. On the posterior clavicle element, they are arranged in rows with their longitudinal axis in line. In the denticle rows on the anterior clavicle element the longitudinal axes of the teeth are aligned parallel to one another.
In position, structure, morphology, size, and size proportion the two ossifications described before agree well with the clavicle elements (serrated appendages) of the Recent Amia calva Linnaeus, Citation1766 (Grande & Bemis, Citation1998:140–141, figs. 87–89). In fossil neopterygians, clavicle elements were made known so far only in rare instances (Bartram, Citation1977; Grande & Bemis, Citation1998). In all cases these elements are of similar appearance. Bartram (Citation1977:216) mentions a specimen of Caturus ? furcatus in which the surface of the serrated appendage “is … bearing small, three-spined denticles,” which correspond to denticle morphology as observed in the clavicle elements of L. gigas. The occurrence of clavicle elements among neopterygians, their morphology, anatomy, function, and homology are discussed by, for example, Bartram (Citation1977), Grande and Bemis (Citation1998), and Liem and Woods (Citation1973).
Note—There is some confusion in the literature concerning the terms “serrated appendages,” “clavicle,” and “clavicle element.” Structures Arratia (Citation2013) named “serrated appendage” in the teleosts Parapholidophorus nybelini (Upper Triassic of Italy) and Leptolepis coryphaenoides (Lower Jurassic of Europe) are different in formation, shape, and position from one another and also from the “serrated appendage” observed by Bartram (Citation1977) in Caturus ? furcatus (Upper Jurassic of Germany). In P. nybelini the “serrated appendage” consists of long, toothed ridges covering the antero-external surface of the cleithrum (Arratia, Citation2013:84, fig. 69). It remains obscure whether these ridges form part of the cleithral surface or whether they are independent ossifications. In L. coryphaenoides the “serrated appendage” is a long and narrow toothed strip of bone, which is attached to the medial margin of the cleithrum (Arratia, Citation2013:125, fig. 100). Also, the teeth on the “serrated appendage” of L. coryphaenoides are unicuspid. In C. ? furcatus occurs a single strip-like “serrated appendage” lying along the upper margin of the rostro-ventral arm of the cleithrum. The external surface of the appendage is covered with three-cuspid denticles being arranged in 14 oblique rows (Bartram, Citation1977:216, fig. 50). Patterson (Citation1977:107, fig. 6) applied the terms “clavicle 1” and “clavicle 2” to two little, multicuspid tooth-bearing bony plates being associated with the external face of the cleithrum in ‘Pholidophorus’ germanicus (Lower Jurassic of Europe). These resemble in shape and position structures on the outer face of the cleithrum of the Recent Amia calva, which were named “clavicle elements (serrated appendages)” by Grande and Bemis (Citation1998) obviously considering the terms “clavicle element” and “serrated appendage” as synonymous. According to Arratia (Citation2013: 23) a “clavicle” in actinopterygians is, however, a bony element articulating with the antero-ventral margin of the cleithrum. It is obvious that all of these structures differ in formation, shape, and position and further investigation is needed to clarify their relationships and homology, which is beyond the scope of this study. Because the structures in question in L. gigas are nearly identical with those of A. calva they are here referred to as “clavicle elements (serrated appendages)” following the terminology of Grande and Bemis (Citation1998).
In summary, specimen FWD_1098 reveals previously unknown skeletal features in L. gigas from the Lower Jurassic of south Germany. These are (1) foramina of different sizes on the maxilla and (2) the formation of two tooth-bearing clavicle elements in the pectoral girdle. The large maxillary foramen may have served for the passage of nerves and vessels supplying either a maxillary pit-line ventral to the infraorbital canal or external cutaneous sense organs such as taste buds in the lip and external skin of the upper jaw. The clavicle elements are morphologically very similar to those present in the living Amia calva but, as in A. calva, specimen FWD_1098 does not yield any hint concerning their function.
DATA AVAILABILITY STATEMENT
The author confirms that the data supporting the findings of this study are available within the article.
DISCLOSURE STATEMENT
No potential conflict of interest was reported by the author.
ACKNOWLEDGMENTS
I thank A. Schmid-Röhl, FWD, for assistance in the collection in her care and for loan of the specimen. G. Arratia and an anonymous reviewer made constructive suggestions for improvement of the manuscript.
LITERATURE CITED
- Agassiz, L. J. R. (1832). Untersuchungen über die fossilen Fische der Lias-Formation. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, 3, 139–149.
- Agassiz, L. J. R. (1833). Recherches sur les Poissons Fossiles (Vol. 2(1), pp. 1–48). Petitpierre.
- Agassiz, L. J. R. (1834). Recherches sur les Poissons Fossiles (Vol. 2, Atlas, pls. 28, 29). Petitpierre.
- Agassiz, L. J. R. (1837). Recherches sur les Poissons Fossiles (Vol. 2(1), pp. 225–264). Petitpierre.
- Allis, E. P. (1889). The anatomy and development of the lateral line system in Amia calva. Journal of Morphology, 2, 463–566. https://doi.org/10.1002/jmor.1050020303
- Allis, E. P. (1897). The cranial muscles and cranial and first spinal nerves in Amia calva. Journal of Morphology, 12, 487–808. https://doi.org/10.1002/jmor.1050120302
- Arratia, G. (2013). Morphology, taxonomy, and phylogeny of Triassic pholidophorid fishes (Actinopterygii, Teleostei). Society of Vertebrate Paleontology, Memoir 13 (Supplement to Journal of Vertebrate Paleontolgy, 33(6)), 1–138. https://doi.org/10.1080/02724634.2013.835642
- Arratia, G., & Huaquín, L. (1995). Morphology of the lateral line system and of the skin of dyplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonner zoologische Monographien, 36, 1–110.
- Bartram, A. W. H. (1975). The holostean fish genus Ophiopsis Agassiz. Zoological Journal of the Linnean Society, 56, 183–205. https://doi.org/10.1111/j.1096-3642.1975.tb00263.x
- Bartram, A. W. H. (1977). The Macrosemiidae, a Mesozoic family of holostean fishes. Bulletin of the British Museum of Natural History (Geology Series), 29(2), 137–234. https://doi.org/10.5962/p.313895
- Blainville, H. D. de (1818). Sur les ichthyolites ou les poissons fossiles. In F. G. Levrault (Ed.), Nouveau dictionnaire d’histoire naturelle, applique´e aux arts, a` l’economie rurale et domestique, a` la Medicine, etc. (pp. 310–395). Deterville.
- Cavin, L., & Suteethorn, V. (2006). A new semionotiform (Actinopterygii, Neopterygii) from Upper Jurassic – Lower Cretaceous deposits of north-east Thailand, with comments on the relationships of semionotiforms. Palaeontology, 49(2), 339–353. https://doi.org/10.1111/j.1475-4983.2006.00539.x
- Coombs, S., Janssen, J., & Webb, J. F. (1988): Diversity of Lateral Line Systems: Evolutionary and Functional Considerations. In F. J. Atema, R. Richard, A. N. Popper, & W. N. Tavolga (Eds.), Sensory Biology of Aquatic Animals (pp. 553–593). Springer.
- Cope, E. D. (1887). Zittel's Manual of Palaeontology. American Naturalist, 21, 1014–1019.
- Devillers, C. (1958). Le système latéral. In P.–P. Grassé (Ed.), Traité de Zoologie (Vol. 13(2), pp. 940–1032). Masson.
- Finger, T. E. (1988): Organization of chemosensory systems within the brain of bony fishes. In F. J. Atema, R. Richard, A. N. Popper, & W. N. Tavolga (Eds.), Sensory Biology of Aquatic Animals (pp. 339–363). Springer.
- Gardiner, B. G. (1984). The relationships of the palaeoniscid fishes, a review based on new specimens of Mimia and Moythomasia from the Upper Devonian of Western Australia. Bulletin of the British Museum (Natural History), Geology series, 37, 173–428.
- Grande, L., & Bemis, W. E. (1998). A Comprehensive Phylogenetic Study of Amiid Fishes (Amiidae) Based on Comparative Skeletal Anatomy. An Emperical Search for Interconnected Patterns of Natural History. Society of Vertebrate Paleontology, Memoir 4 (Supplement to Journal of Vertebrate Paleontolgy, 18), 1–696. https://doi.org/10.1080/02724634.1998.10011114
- Hay, O. P. (1929). Second bibliography and catalogue of the fossil Vertebrata of North America. Publications of the Carnegie Institute of Washington, 390, 1–2003.
- International Commission on Zoological Nomenclature (1999). International Code of Zoological Nomenclature (4th ed.). The International Trust for Zoological Nomenclature.
- Jaekel, O. (1929). Lepidotus und Leptolepis aus dem oberen Lias von Dobbertin, Mecklenburg. Mitteilungen der Mecklenburgischen Geologischen Landesanstalt, XXXVIII (Neue Folge III), 15–25.
- Jakubowski, M. (1966). Cutaneous sense organs of fishes. IV. The lateral-line organs in the perch-like Lucioperca lucioperca and the perch Perca fluviatilis, their topography, innervation, vascularization, and structure. Acta Biologica Cracoviensia, Series Zoologia, 9, 137–149.
- Jakubowski, M. (1967). Cutaneous sense organs of fishes. VI. The structure, topography, and innervation of lateral line organ in the burbot (Lota lota). Acta Biologica Cracoviensia, Series Zoologia, 10, 39–47.
- Kindred, J. E. (1919). The skull of Amiurus. Illinois Biological Monographs, 5(1), 1–120.
- Lane, J. A., & Ebert, M. (2015). A taxonomic reassessment of Ophiopsis (Halecomorphi, Ionoscopiformes). With a revision of Upper Jurassic species from the Solnhofen Archipelago, and a new genus of Ophiopsidae. Journal of Vertebrate Paleontology 35, e883238. https://doi.org/10.1080/02724634.2014.883238.
- Lehman, J.–P. (1966). Actinopterygii. In J. Piveteau (Ed.): Traité de Paléontologie: Actinoptérygiens, Dipneustes, Crossoptérygiens (Vol. IV(3), pp. 1–242). Masson.
- Lekander, B. (1949). The sensory line system and the canal bones in the head of some Ostariophysi. Acta Zoologica, 30, 1–131. https://doi.org/10.1111/j.1463-6395.1949.tb00503.x
- Liem, K. F., & Woods, L. P. (1973). A probable homologue of the clavicle in the holostean fish Amia calva. Journal of Zoology, 170, 521–531. https://doi.org/10.1111/j.1469-7998.1973.tb05067.x
- Linnaeus, C. (1766). Systema Naturae (12th rev. ed., Vol. 1). Impensis Direct Laurentii Salvii.
- López-Arbarello, A. (2012). Phylgenetic Interrelationships of Ginglymodian Fishes (Actinopterygii: Neopterygii). PLoS ONE 7(7), e39370, https://doi.org/10.1371/journaL.pone.0039370
- Leuzinger, L., Cavin, L., López-Arbarello, A., & Billon-Bruyat, J.–P. (2020). Peculiar tooth renewal in a Jurassic ray-finned fish (Lepisosteiformes, †Scheenstia sp.). Palaeontology, 63(1), 117–129. https://doi.org/10.1111/pala.12446
- Patterson, C. (1977). Cartilage bones, dermal bones and membrane bones, or the exoskeleton versus the endoskeleton. In S. M. Andrews, R. S. Miles & A. D. Walker (Eds.), Problems in Vertebrate Evolution (Linnean Society Symposium Series No. 4, pp. 77–121). Linnean Society London.
- Pehrson, T. (1944). The development of latero-sensory canal bones in the skull of Esox lucius. Acta Zoologica, 25, 135–157. https://doi.org/10.1111/j.1463-6395.1944.tb00350.x
- Quenstedt, F. A. (1847). Anzeige der akademischen Feier des Geburtsfestes Seiner Majestät des Königs Wilhelm von Württemberg im Namen des Rektors und des Senats der königlichen Eberhard-Karls-Universität zu Tübingen, nebst Lepidotus im Lias Epsilon Württembergs. L. F. Fues.
- Quenstedt, F. A. (1852). Handbuch der Petrefactenkunde. Laupp.
- Quenstedt, F. A. (1858). Der Jura. Laupp & Siebeck.
- Rixon, A. E. (1976). Fossil Animal Remains. Their Preparation and Conservation. Athlone Press.
- Rutzky, I. S., Elvers, W. B., Maisey, J. G., & Kellner, A. W. A. (1995). Chemical preparation techniques. In P. Leiggi & P. May (Eds.), Vertebrate Paleontological Techniques (Vol 1, pp. 155–186). Cambridge University Press.
- Schultze, H.–P. (2008). Nomenclature and homologization of cranial bones in actinopterygians. In G. Arratia, H.–P. Schultze, & M. V. H. Wilson (Eds.), Mesozoic Fishes 4 – Homology and Phylogeny: Proceedings of the international meeting, Miraflores de la Sierra, 2005 (pp. 23–48). F. Pfeil.
- Stensiö, E. A. (1947). The sensory lines and dermal bones of the cheek in fishes and amphibians. Kungliga Svenska Vetenskapsakademien Handlingar, Serie 3, 24, 1–195.
- Thies, D. (1989a). Lepidotes gloriae, sp. nov. (Actinopterygii: Semionotiformes) from the Late Jurassic of Cuba. Journal of Vertebrate Paleontology, 9, 18–40. https://doi.org/10.1080/02724634.1989.10011736
- Thies, D. (1989b). Sinneslinien bei dem Knochenfisch Lepidotes elvensis (Blainville, 1818) (Actinopterygii, †Semionotiformes) aus dem Oberlias (Unter-Toarcium) von Grimmen in der DDR. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 1989, 692–704. https://doi.org/10.1127/njgpm/1989/1989/692
- Thies, D. (1989c). Cranial sensory lines in Jurassic semionotiform fishes (Actinopterygii) [Abstract]. Journal of Vertebrate Paleontology, 9(Suppl. to no. 3), 41A.
- Thies, D. (1991). The osteology of the bony fish Tetragonolepis semicincta Bronn 1830 (Actinopterygii, †Semionotiformes) from the Early Jurassic (Lower Toarcian) of Germany. Geologica et Palaeontologica, 25, 251–297.
- Thies, D., Stevens, K., & Ansorge, J. (in press). A new lepisosteiform neopterygian (Actinopterygii) from the lower Toarcian Grimmen Formation (Lower Jurassic) of Mecklenburg and Western Pomerania, Germany. Paläontologische Zeitschrift.
- Thies, D., Stevens, K., & Stumpf, S. (2019). Stomach contents of the Early Jurassic fish †Lepidotes Agassiz, 1832 (Actinopterygii, Lepisosteiformes) and their palaeoecological implications. Historical Biology 33, 868–879. https://doi.org/10.1080/08912963.2019.1665040
- Woodward, A. S. (1895). Catalogue of the Fossil Fishes in the British Museum (Natural History) (Part III). Trustees of the British Museum.
- Woodward, A. S. (1897). On the Fossil Fishes of the Upper Lias of Whitby: Part III. Proceedings of the Yorkshire Geological and Polytechnical Society, 13(3), 325–337. https://doi.org/10.1144/pygs.13.3.325