
Abstract
Carbapenem-resistant Salmonella enterica (S. enterica) pose a significant threat to public health, causing gastroenteritis and invasive infections. We report the first emergence of a carbapenem-resistant S. enterica serovar London strain, A132, carrying the blaNDM-5 gene in China. Whole-genome sequencing and bioinformatics analysis assigned A132 to be ST155, a multidrug-resistant clone frequently reported in China. The strain A132 exhibited resistance to multiple antibiotics, with 20 acquired antibiotic resistance genes (ARGs) identified, predominantly located on the IncFIB plasmid (pA132-1-NDM). Notably, the blaNDM-5 gene was located within an IS26 flanked-class 1 integron-ISCR1 complex, comprising two genetic cassettes. One cassette is the class 1 integron, which may facilitate the transmission of the entire complex, while the other is the blaNDM-5-containing ISCR1-IS26-flanked cassette, carrying multiple other ARGs. Genbank database search based on the blaNDM-5-carrying cassette identified a similar genetic context found in transmissible IncFIA plasmids from Escherichia coli (p91) and Enterobacter hormaechei (p388) with a shared host range, suggesting the potential for cross-species transmission of blaNDM-5. To our knowledge, this is the first reported case of Salmonella serovar London ST155 harboring blaNDM-5 gene. Phylogenetic analysis indicated a close relationship between A132 and eight S. London ST155 strains isolated from the same province. However, A132 differed by carrying the blaNDM-5 gene and four unique ARGs. Given the high transmissibility of the F-type plasmid harboring blaNDM-5 and 18 other ARGs, it is imperative to implement vigilant surveillance and adopt appropriate infection control measures to mitigate the threat to public health.
Introduction
Salmonella is a major causative agent of gastroenteritis on a global scale.Citation1 Annually, non-typhoidal Salmonella (NTS) is responsible for an estimated 200 million to over 1 billion infections globally, resulting in 93 million cases of gastroenteritis and 155,000 fatalities.Citation2,Citation3 Although NTS typically induces self-limiting gastroenteritis, invasive NTS infections frequently result in substantial mortality rates.Citation4 In 2017, NTS was estimated to cause 535,000 invasive infections, contributing to 77,500 deaths worldwide.Citation5
The primary treatment for invasive salmonellosis previously relied on chloramphenicol, amoxicillin (ampicillin), and cotrimoxazole (trimethoprim/sulfamethoxazole). However, widespread resistance to these first-line drugs has prompted the endorsement of alternative therapeutic strategies, encompassing extended-spectrum cephalosporins (eg, ceftriaxone) and fluoroquinolones (eg, ciprofloxacin).Citation6 Unfortunately, the extensive use of these secondary antimicrobials has led to the development of resistance to extended-spectrum cephalosporins and fluoroquinolones in NTS strains, presenting a substantial hurdle in the medical management of infections.Citation6 Carbapenems and azithromycin are currently considered as ultimate therapeutic alternatives for treating invasive Salmonella infections induced by multidrug and extensively drug-resistant strains.Citation7
Carbapenemase-producing Enterobacteriaceae pose a growing threat to public health due to their resistance to carbapenems, a crucial class of first-line antibiotics used in the treatment of multidrug-resistant (MDR) bacterial infections.Citation8 Although carbapenem resistance remains rare in NTS, several studies have documented the presence of carbapenemase-producing genes in Salmonella, including variants such as blaKPC-1, blaNDM-1 and blaVIM-1.Citation9,Citation10
In this study, we report the identification of an S. enterica serovar London strain carrying the blaNDM-5 gene in China. To the best of our knowledge, this represents the first documentation of blaNDM-5 in S. London. Whole-genome sequencing (WGS) was employed to elucidate the comprehensive genomic profile, coupled with extensive bioinformatics analyses, to enhance our understanding of the global dissemination of NDM.
Materials and Methods
Bacterial Isolation and Identification
The blaNDM-5-carrying S. enterica serovar London strain A132 was collected once from the stool sample of a 35-year-old female patient in the First People’s Hospital of Huzhou on 21st July 2023. The patient suffered from severe diarrhea, which progressed from 3 to 4 times a day before treatment to about 10 times on the day of diagnosis. The routine blood test showed that the patient’s WBC (9.4 × 10^9/L), neutrophil ratio 82.5%, and high-sensitivity C-reactive protein (15.29 mg/L) were all beyond the normal range, suggesting a bacterial infection. This patient was hospitalized due to serious fever and diarrhea. The species of A132 was identified using the Matrix-Assisted Laser Desorption Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS).Citation11
Antimicrobial Susceptibility Testing
The strain was cultured on LB agar plates. One hundred and forty-five microliter bacterial suspension of 0.5-McFarland turbidity was mixed with 3mL 0.45% NaCl solution. The AST-GN13 card filled with the mixture was used. The minimum inhibitory concentrations (MICs) of commonly used antibiotics, including ampicillin, amoxicillin/clavulanic acid, piperacillin, cefazolin, ceftazidime, ceftriaxone, cefepime, aztreonam, imipenem, meropenem, amikacin, gentamicin, ciprofloxacin, levofloxacin, tetracycline, nitrofurantoin and sulfamethoxazole/trimethoprim, were determined by the Vitek2 compact system (BioMérieux, France) following the manufacturer’s instructions. Antimicrobial sensitivity results were interpreted according to the Clinical and Laboratory Standards Institute guidelines (M100-S32).Citation12 The Escherichia coli ATCC25922 was used as quality control and negative control for the antimicrobial susceptibility tests.
Whole-Genome Sequencing
Genomic DNA was extracted using Wizard® Genomic DNA Purification Kit (Promega, Madison, WI) according to the manufacturer’s protocol. WGS was performed using both the PacBio Sequel II platform (Pacific Biosciences, Menlo Park, CA, USA) with the SMRT bell TM Template kit (Pacific Biosciences) and the Illumina NovaSeq platform (San Diego, CA, USA) with a TruSeq Nano library kit (Illumina) according to the manufacturer’s instructions.
Bioinformatics Analysis
The long and short WGS reads were trimmed with filtlong (https://github.com/rrwick/Filtlong) and fastp, respectively.Citation13 Hybrid assembly was performed based on trimmed long and short reads using Unicycler v0.5.0 with default settings, resulting in a complete genome assembly.Citation14 WGS-based Salmonella serotyping was performed using SeqSero with default settings.Citation15 The obtained sequences were submitted to PubMLST database (https://pubmlst.org/) to determine the allele number and specific sequence type (ST). MOB-suite v3.1.4 was adopted to predict plasmid sequences from the assembly genome and identify their replicon types, mobility and host range.Citation16
Gene predictions and functional annotations were performed with RAST server.Citation17 The presence of acquired antibiotic resistance genes (ARGs) and chromosomal resistance mutations was detected with ResFinder v4.1.Citation18 Virulence factors (VFs) were screened using the VFDB database.Citation19 The search for insertion sequence (IS) elements and their characterization down to the family was carried out correspondingly using digIS and ISfinder.Citation20,Citation21 IntegronFinder v2.0 was used to detect complete integrons.Citation22 Salmonella pathogenicity islands (SPIs) were identified with SPIFinder.Citation23
Core genome single nucleotide polymorphisms (cgSNPs) were extracted with Parsnp v1.2, using the A132 complete genome as a reference.Citation24 Recombination sites were removed using Gubbins v3.3 with default parameters and a starting tree created by Parsnp using a GTR substitution model.Citation25 Pairwise SNP distances were calculated with SNP-sites.Citation26 The maximum likelihood phylogeny based on the concatenated data was inferred by IQ-TREE 2 with a HKY model.Citation27 The best topology was assessed by 1000 ultrafast bootstraps. The phylogenetic tree was visualized with the interactive Tree of Life (iTOL) web application.Citation28 Plasmid alignment was generated using BRIG v0.95.Citation29
Results
Phenotypes and Genotypes of S. London A132
As summarized in , S. London A132 exhibited sensitivity solely to aztreonam, while demonstrating resistance to ampicillin, ampicillin/sulbactam, cefepime, ceftriaxone, ceftazidime, ciprofloxacin, ertapenem, imipenem, piperacillin/tazobactam, and trimethoprim/sulfamethoxazole, so-called multidrug resistance (MDR). Additionally, an intermediate resistance pattern was observed with levofloxacin.
Table 1 Antibiotic Susceptibilities of S. London A132
Based on complete genome assembly using both third- and second-generation sequencing, strain A132 was found to be composed of one chromosome (4597.8 kb) and two plasmids—IncFIB-type (111.9 kb) and IncQ1-type (6.1 kb). In silico typing affiliated A132 to serotype London with an antigenic profile: 3.10: l, v:1,6 (O antigen: 3, 10, H antigen Phase 1: v, H antigen Phase 2: 1.6) based on the Kauffmann-White scheme. Additionally, according to Achtman’s MLST scheme, S. London strain A132 was classified as ST155.
Antimicrobial Resistance and Virulence-Associated Genes
ResFinder-based screening identified 20 acquired ARGs () that conferred resistance to β-lactams (blaNDM-5, blaTEM-1), aminoglycoside [aac(6’)-Iaa, aac(3)-IId, aadA16, aph(6)-Id, aph(3”)-Ib, aadA2, aac(6’)-Ib-cr], quinolone [aac(6’)-Ib-cr, qnrB6), sulfonamide (three copies of sul1, sul2), diaminopyrimidine (dfrA12, dfrA27), macrolide (mphA), tetracycline (tetA), chloramphenicol (floR) and rifamycin (ARR-3). Moreover, a mutation in parC (Trp57→Ser) was identified with the absence of gyrA mutation, which explained the low ciprofloxacin MICs.Citation30
Table 2 Genetic Mechanisms of Multidrug Resistance of S. London A132
Using SPIFinder, A132 was found to harbor SPI-1 to SPI-5, SPI-9, and SP-12 to SP-14 on its chromosome. The VFDB led to the detection of 154 VFs (Supplementary Table S1), most of which were associated to bacterial secretion system (38% of the VFs detected, eg hilA, invB, sopA) and fimbrial adherence determinants (27% of the VFs detected, eg Agf, Bcf, Fim). Several VFs related to the evasion of the immune system (ie macrophage inducible genes (mig-14)), magnesium uptake (mgtB, mgtC), stress adaptation (sodCI), toxin secretion (spvB, cdtB, pltA, pltB) and autotransporters (ehaB) were also encoded in the genome. All of the VFs were located on the chromosome.
Characterization of the blaNDM-5 Carrying Plasmid in A132
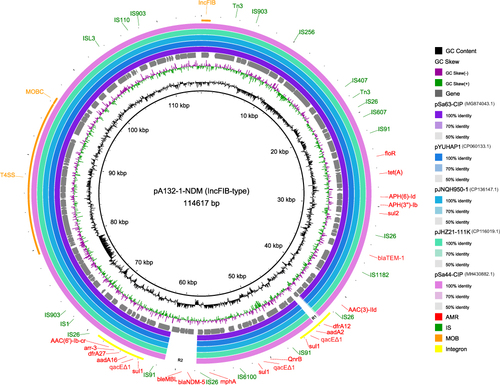
The blaNDM-5 gene was located on the IncFIB-type plasmid, herein designated pA132-1-NDM. Notably, pA132-1-NDM carried 95% (21/22) of the acquired ARGs. According to MOB-suite, pA132-1-NDM was predicted to be conjugative due to the presence of relaxase and mate-pair formation markers. However, the origin of the transfer (oriT) region was not detected. RAST annotation revealed the presence of genes associated with membrane transport (eg virB, virC, virD, pinF), DNA metabolism (eg hsdM, hsdR, hsdS, rhuM), protein folding (eg dsbA, dsbB, dsbC, dsbD), respiration (eg cchl, hyp5, ccmA) and transposable elements integrons (int, intI1, intI2) were also identified in pA132-1-NDM. A total of 21 insertion sequences were detected, including IS1, IS3, IS5, IS6, IS110, IS1182, IS256, IS91, ISL3 and Tn3 families. Moreover, pA132-1-NDM carried two complete class 1 integrons with the common qacE∆1-sul1 region, conferring resistance to quaternary ammonium salts and sulfonamides. These integrons integrated resistance gene cassettes, including aac(6’)-Ib-cr-aar3-dfrA27-aadA16 and dfrA12-aadA2 gene arrays, respectively.
A megaBLAST of the pA132-1-NDM sequence retrieved five highly similar IncFIB-type plasmids from previously reported Salmonella strains (approximately 96% identity and 100% coverage). Information about these plasmids, which all isolated in China between 2013 and 2022, is given in Supplementary Table S2. The serotypes of these strains were London (n=4) and Thompson (n = 1), and the sources were swine (n = 2), food (n = 2) and human (n = 1). According to plasmid genome alignment, all of these plasmids were missing two regions in comparison to pA132-1-NDM (). The two regions were 11.7 kb apart, with the first region (R1, 1.7 kb) contained only the dfrA12 gene, while the second region (R2, 3.6 kb) comprised blaNDM-5 and three other genes (bleMBL, trpF, and dsbD), implying two independent horizontal gene transfer events.
Figure 1 Genetic map of pA132-1-NDM with five other homologous IncFIB-type plasmids in Salmonella. Concentric rings represent the similarity between the reference sequence (pA132-1-NDM) in the inner ring and other sequences in the outer rings. The resistance and IS genes are annotated in red or green fonts, respectively. The sequence related to plasmid mobility and integron were highlighted with Orange or yellow rings, correspondingly.
Comparative Analysis of the Genetic Context of blaNDM-5
Detailed examination of the genetic context of blaNDM-5 showed that it is located within a region flanked by two IS26 sequences, about 13 kb in length, which composed of two gene cassettes linked by an ISCR1 (IS91 family) element (). One of the cassettes is the so-called class 1 integron cassette (IS26-intI1-Ib-cr-aac(6’)-arr-3-dfrA27-aadA16-qacEΔ1-sul1) which comprised of an intI1 gene truncated by one of the IS26 copies, and the aac(6’)-Ib-cr-arr-3-dfrA27-aadA16-qacEΔ1-sul1 ARGs cassette. The other cassette is named ISCR1-IS26-flanked cassette (ISCR1-dsbd-trpF-bleMBL-blaNDM-5-IS26) in our study, with genes ISCR1, dsbd (Dsbd superfamily protein), trpF (phosphor ribosyl anthranilate isomerase) and bleMBL (bleomycin resistance gene) located upstream, and an IS26 downstream. Together, the two cassettes form the so-called class 1 integron-ISCR1 complex which is commonly found in gram-negative bacteria.Citation31
Figure 2 Schematic representation of the blaNDM-5-containing class 1 integron-ISCR1 complex found in the A132 and other two representative sequences identified in E. coli plasmid p91_NDM (MN007141.1) and Ent. hormaechei plasmid p388 (CP021168.1) from Genbank. This image was built using the R package gggenomes (https://github.com/thackl/gggenomes).
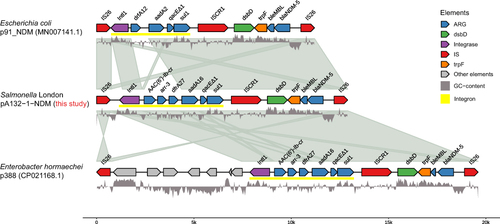
To investigate the potential origin of blaNDM-5-containing cassette in A132 strain, we carried out a series of NCBI BLAST search using nucleotide sequences. We found that the ISCR1-IS26-flanked cassette (ISCR1-dsbd-trpF-bleMBL-blaNDM-5-IS26) of pA132-1-NDM showed a perfect match with a previously reported blaNDM-5 harboring plasmid p91_NDM of E. coli (MN007141.1) with 100% identity and 100% coverage. Moreover, the BLAST search using the entire class 1 integron-ISCR1 complex sequence in pA132-1-NDM yielded the one identified in plasmid p388 from Enterobacter hormaechei (CP021168.1) which exhibited a high degree of similarity (100% identity and 99% coverage). The sequence alignment of the class 1 integron-ISCR1 complex in A132 strain with the two matched results from BLAST search (p91_NDM_Escherichia coli and p388_Enterobacter hormaechei) was shown in . Interestingly, both p91_NDM and p388 were identified as conjugative IncFIA-type plasmids with a broad host range among Enterobacterales, while the pA132-1-NDM found in our study was identified as IncFIB-type plasmid.
Phylogenetic Analysis of Chinese S. London ST155 Isolates
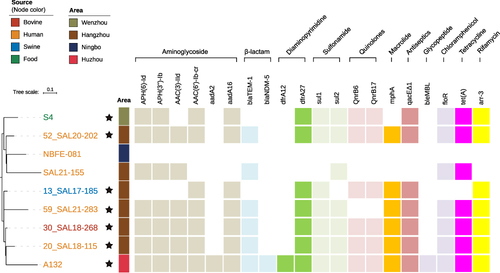
To delineate genetic distinctions between A132 and other S. London ST155 strains in China, we retrieved 195 additional Chinese S. London ST155 strains from the EnteroBase database (up until December 2023).Citation32 A cgSNP analysis revealed differences ranging from 14 to 169 SNPs between these strains and A132 (Supplementarys Figure S1 and Table S3). Of the 164 strains with identifiable sampling sources, the overwhelming majority originated from humans (56.7%, 93/164), swine (24.4%, 30/164) and food (14.6%, 24/164). We further narrowed down to eight closely related strains using a threshold of 40 cgSNPs ( and Supplementary Table S3).Citation33 These strains were isolated from three different cities in Zhejiang Province between 2017 and 2021, including Hangzhou (n = 6), Ningbo (n = 1), and Wenzhou (n = 1). Plasmid detection through MOB-recon showed that except for NBFE-081 and 13_SAL17-185, the other six strains all carried an IncFIB-type plasmid. Apart from the strain NBFE-081 from Ningbo, the other seven strains were predicted to be MDR due to the presence of multiple common ARGs. Interestingly, compared with these closely related strains, the A132 strain in our study carried four unique ARGs, including aadA2, dfrA12, bleMBL and blaNDM-5 which all located within the class 1 integron-ISCR1 complex.
Figure 3 Phylogenetic association between A132 and its eight closely related Chinese S. London ST155 isolates using cgSNP strategy. Antimicrobial resistant genes were categorized into groups based on their resistance to different types of antibiotics, as shown in boxes with different colors. The absence of box indicates the absence of particular resistance gene in that strain. The source of each isolate is shown as colored terminal nodes. Different colored strips indicate the city where the strains were isolated. The black star indicates the presence of an IncFIB plasmid in that strain.
Discussion
NDM-5 carbapenemase was primarily detected in E. coli and remains relatively rare in other members of the Enterobacteriaceae family, such as NTS and Klebsiella pneumoniae.Citation34,Citation35 However, our study reveals the emergence of a Salmonella strain, designated as A132, harboring the NDM-5 carbapenemase gene. A132 belongs to the serovar London, which recognized as one of the top five prevalent serovars responsible for human salmonellosis in China.Citation36 Furthermore, the sequence type of A132 is classified as ST155, an MDR clone that has been frequently reported in China, thus posing a significant threat to public health.Citation37 A132 exhibited multidrug resistance and carried a total of 20 acquired ARGs. These ARGs were in concordance with the DST phenotypes, with 95% of the ARGs located on the IncFIB plasmid (pA132-1-NDM). Previous reports in Salmonella have mainly identified blaNDM genes on IncX3 and IncFII plasmids.Citation38–40 Notably, our study represents the first documentation of blaNDM-5 located on a transmissible IncFIB plasmid of Salmonella. IncFIB plasmids, often referred to as ColV plasmids, have predominantly been associated with serovars Kentucky, Typhimurium, Heidelberg and Schwarzengrund.Citation41–43 The results of plasmid BLAST search in Genbank database retrieved five highly homologous IncFIB plasmids in serovar London and Thompson, all isolated from China. Several studies have suggested that IncFIB plasmids likely contribute to increased colonization in the cecum of poultry, providing an explanation for their persistence in food animal populations.Citation42,Citation43 These findings suggest that IncFIB plasmids may potentially contribute to the dissemination of ARGs among bacteria in various livestock species.
Based on the comparative analysis of the blaNDM-5 gene context in strain A132 with those in public databases using NCBI BLAST search (), we identified the ISCR1-IS26-flanked cassette (ISCR1-dsbd-trpF-bleMBL-blaNDM-5-IS26) as a highly conserved and stable domain in the IncF-type plasmids of E.coli and Enterobacter hormaechei. Additionally, previous studies have reported blaNDM-5 gene in IncX3-type plasmids of Enterobacteriaceae, located in the roughly similar genetic context (IS3000-ISAba125Δ-IS5-ΔISAba125-blaNDM-5-bleMBL-trpF-dsbC-cutA-IS26), with the blaNDM-5 gene flanked by IS elements and located adjacent to three other ARGs (bleMBL-trpF-dsbC).Citation39,Citation40,Citation44 Despite the presence or absence of other gene elements in the genetic context (such as ΔISAba125 and cutA which were not found in pA132-1-NDM), the combination of four ARGs (blaNDM-5-bleMBL-trpF-dsb) were found to be conserved across all blaNDM-5-harboring IncX3- and IncF-type plasmids reported thus far. These evidences together emphasize the conservation and significance of the four ARGs as a basic unit in the transmission and expression of carbapenemase, suggesting a potential pattern for the dissemination of blaNDM-5 gene.Citation40,Citation45
Moreover, the location of blaNDM-5 gene in the vicinity of a class 1 integron seems to enhance its dissemination. The blaNDM-5-containing ISCR1-IS26-flanked cassette in pA132-1-NDM was found to be linked to a complete class 1 integron via transposition of an ISCR1 element. It was known that Class 1 integrons are widely distributed in the environmental microbial genomes and are implicated in the dissemination of many ARGs.Citation46 According to the model proposed by Toleman et al, the presence of ISCR1 can mediate the fusion of class 1 integron with other IS-flanked ARGs-carrying cassettes, forming a class 1 integron-ISCR1 complex.Citation31 Interestingly, the five highly similar IncFIB-type plasmids in Salmonella strains retrieved from the megaBLAST search all showed the same pattern of the blaNDM-5-carrying cassette missing in comparison to pA132-1-NDM (). Therefore, we suspect that the presence of blaNDM-5 gene in A132 may be the result of recent integration of the ISCR1-IS26-flanked cassette mediated by the ISCR1 element, leading to the formation of a class 1 integron-ISCR1 complex, as demonstrated by Toleman et al.Citation31 In support of our speculation, a BLAST search using the class 1 integron-ISCR1 complex of p132-1-NDM revealed bundles of highly identical genetic structures in IncFIA plasmids of E. coli and Ent. hormaechei. These evidence collectively suggest that the blaNDM-5-carrying cassette is frequently transferred as an integral complex with class 1 integron rather than as a standalone gene cassette, as also implicated in previous studies with E. coli.Citation47,Citation48 Nevertheless, future research is necessary to validate the role of class 1 integron and ISCR1 element in the dissemination of blaNDM-5 gene.
Using cgSNP-based phylogenetic analysis, we found that A132 shares a close genomic relationship with numerous Chinese S. London ST155 strains retrieved from EnteroBase. These prevalent Chinese S. London ST155 isolates have been predominantly sourced from humans, swine, and food products, suggesting that S. London infections in humans often originate from zoonotic transmission.Citation49 Therefore, it is highly probable that the patient in our study had a foodborne infection. In addition, recent studies have reported a rising prevalence of S. London isolates associated with foodborne human salmonellosis in Zhejiang Province.Citation50 The identification of eight isolates in Zhejiang Province, differing from A132 by a threshold of 40 cgSNPs, yet carrying similar plasmids and ARGs, underscores the necessity for vigilant surveillance of S. London pathogens.Citation33 Notably, A132 possesses four unique ARGs (blaNDM-5,bleMBL,trpF and dsbd genes) compared to these strains. This suggests a recent acquisition of the blaNDM-5 gene and the other four unique ARGs by A132 from the environment, possibly through horizontal gene transfer mechanisms. This acquisition further amplifies the multidrug resistance profile of Salmonella, presenting a formidable clinical challenge in combating antibiotic resistance.
From One Health perspective, the emergence of a blaNDM-5-carrying S. enterica serovar London strain in China presents significant clinical implications.Citation51 Given the pivotal role of blaNDM-5 gene in multi-drug resistance, a preliminary search on PubMed indicates a steady increase in reported cases of blaNDM-5 over the past decades, possibly linked to the excessive usage of antibiotics. While E. coli accounts for the majority of reported cases, the presence of blaNDM-5 gene in Salmonella, as demonstrated in our study, which is located alongside other ARGs in proximity to IS elements and housed within a highly transmissible IncFIB plasmid adds complexity to the management of infections. Therefore, sustained attention needs to be warranted, focusing on genotyping, plasmid characterization, and phylogenetic analysis to better understand the transmission dynamics of blaNDM-5-carrying Salmonella, guiding public health interventions.
Conclusion
Our study documented the first isolation of an NDM-5-producing Salmonella enterica serovar London strain in China. The emergence of the blaNDM-5 gene in a clinical MDR S. London ST155 strain represents a significant threat to public health. Through EnteroBase database search, we retrieved eight highly similar Chinese S. London ST155 strains from other cities in Zhejiang Province, which mostly exhibited MDR, highlighting the importance of the surveillance for S. London ST155. We suspect that the existence of blaNDM-5 gene within an ISCR1-IS26-flanked cassette on the IncFIB plasmid is likely a result of recent horizontal gene transfer from other Enterobacteriaceae. The blaNDM-5 gene carried by a transmissible IncFIB plasmid may cause an increased risk of horizontal transfer of the blaNDM-5 gene. Moreover, the class 1 integron and ISCR1 element in the vicinity of blaNDM-5 gene may also benefit the mobilization and dissemination of blaNDM-5, which highlights the necessity for implementing proactive clinical controls to mitigate the spread of blaNDM-5 and its associated multidrug resistance.
Data Sharing Statement
The datasets presented in this study can be found in online repositories. The genome sequence of the strain has been deposited in the NCBI database under BioProject accession number PRJNA1057448.
Ethics Approval and Informed Consent
The authors certify that the patient consent form has been obtained. Written informed consent was obtained from the patient for publication of this Case report and any accompanying images. The studies involving human participants were reviewed and approved by the Ethics Committee of the First People’s Hospital of Huzhou.
Disclosure
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Funding
References
- Duff N, Steele AD, Garrett D. Global action for local impact: the 11th international conference on typhoid and other invasive salmonelloses. Clin Infect Dis. 2020;71(Suppl 2):S59–S63. doi:10.1093/cid/ciaa236
- Chlebicz A, Śliżewska K. Campylobacteriosis, salmonellosis, yersiniosis, and listeriosis as zoonotic foodborne diseases: a review. Int J Environ Res Public Health. 2018;15(5):863. doi:10.3390/ijerph15050863
- Castro-Vargas RE, Herrera-Sánchez MP, Rodríguez-Hernández R, Rondón-Barragán IS. Antibiotic resistance in Salmonella spp. isolated from poultry: a global overview. Vet World. 2020;13(10):2070. doi:10.14202/vetworld.2020.2070-2084
- Foley SL, Lynne AM. Food animal-associated Salmonella challenges: pathogenicity and antimicrobial resistance. J Anim Sci. 2008;86(suppl_14):E173–E187. doi:10.2527/jas.2007-0447
- Stanaway JD, Parisi A, Sarkar K, et al. The global burden of non-typhoidal salmonella invasive disease: a systematic analysis for the global burden of disease study 2017. Lancet Infect Dis. 2019;19(12):1312–1324. doi:10.1016/S1473-3099(19)30418-9
- Crump JA, Sjölund-Karlsson M, Gordon MA, Parry CM. Epidemiology, clinical presentation, laboratory diagnosis, antimicrobial resistance, and antimicrobial management of invasive Salmonella infections. Clin Microbiol Rev. 2015;28(4):901–937. doi:10.1128/CMR.00002-15
- McKenna M. Antibiotic resistance: the last resort. Nature. 2013;499(7459):394–396. doi:10.1038/499394a
- Suay-García B, Pérez-Gracia MT. Present and future of carbapenem-resistant Enterobacteriaceae (CRE) infections. Antibiotics. 2019,August 19;8: 3 122 doi:10.3390/antibiotics8030122
- Fernández J, Guerra B, Rodicio M. Resistance to carbapenems in Non-Typhoidal Salmonella enterica serovars from humans, animals and food. Veterinary Sciences. 2018;5(2):40. doi:10.3390/vetsci5020040
- Nordmann P, Dortet L, Poirel L. Carbapenem resistance in Enterobacteriaceae: here is the storm! Trends Mol Med. 2012;18(5):263–272. doi:10.1016/j.molmed.2012.03.003
- Sogawa K, Watanabe M, Sato K, et al. Use of the MALDI BioTyper system with MALDI–TOF mass spectrometry for rapid identification of microorganisms. Anal Bioanal Chem. 2011;400:1905–1911. doi:10.1007/s00216-011-4877-7
- Humphries R, Bobenchik AM, Hindler JA, Schuetz AN. Overview of changes to the clinical and laboratory standards institute performance standards for antimicrobial susceptibility testing, M100. J Clin Microbiol. 2021;59(12):10–1128. doi:10.1128/JCM.00213-21
- Chen S, Zhou Y, Chen Y, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34(17):i884–i890. doi:10.1093/bioinformatics/bty560
- Wick RR, Judd LM, Gorrie CL, Holt KE. Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Computational Biology. 2017;13(6):e1005595. doi:10.1371/journal.pcbi.1005595
- Zhang S, Den Bakker HC, Li S, et al. SeqSero2: rapid and Improved Salmonella serotype determination using whole-genome sequencing data. Appl Environ Microbiol. 2019;85(23):e01746–19. doi:10.1128/AEM.01746-19
- Robertson J, Nash JH. MOB-suite: software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microbial Genomics. 2018;4(8). doi:10.1099/mgen.0.000206
- Aziz RK, Bartels D, Best AA, et al. The RAST server: rapid annotations using subsystems technology. BMC Genomics. 2008;9(1):75. doi:10.1186/1471-2164-9-75
- Florensa AF, Kaas RS, Clausen PTLC, Aytan-Aktug D, Aarestrup FM. ResFinder–an open online resource for identification of antimicrobial resistance genes in next-generation sequencing data and prediction of phenotypes from genotypes. Microbial Genomics. 2022;8(1). doi:10.1099/mgen.0.000748
- Liu B, Zheng D, Zhou S, Chen L, Yang J. VFDB 2022: a general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022;50(D1):1 doi:10.1093/nar/gkab1107
- Puterová J, Martínek T. digIS: towards detecting distant and putative novel insertion sequence elements in prokaryotic genomes. BMC Bioinf. 2021;22(1):258. doi:10.1186/s12859-021-04177-6
- Siguier P, Pérochon J, Lestrade L, Mahillon J, Chandler M. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006;34(suppl_1):D32–D36. doi:10.1093/nar/gkj014
- Néron B, Littner E, Haudiquet M, Perrin A, Cury J, Rocha EP. IntegronFinder 2.0: identification and analysis of integrons across bacteria, with a focus on antibiotic resistance in Klebsiella. Microorganisms. 2022;10(4):700. doi:10.3390/microorganisms10040700
- Roer L, Hendriksen RS, Leekitcharoenphon P, et al. Is the evolution of Salmonella enterica subsp. enterica linked to restriction-modification systems? Msystems. 2016;1(3):10–1128. doi:10.1128/mSystems.00009-16
- Treangen TJ, Ondov BD, Koren S, Phillippy AM. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014;15(11):524. doi:10.1186/s13059-014-0524-x
- Croucher NJ, Page AJ, Connor TR, et al. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015;43(3):e15–e15. doi:10.1093/nar/gku1196
- Page AJ, Taylor B, Delaney AJ, et al. SNP-sites: rapid efficient extraction of SNPs from multi-FASTA alignments. Microbial Genomics. 2016;2(4). doi:10.1099/mgen.0.000056
- Nguyen LT, Schmidt HA, von Haeseler A, Minh BQ. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Molecul Biol Evolu. 2015;32(1):268–274. doi:10.1093/molbev/msu300
- Letunic I, Bork P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021;49(W1):W293–W296. doi:10.1093/nar/gkab301
- Alikhan NF, Petty NK, Zakour NLB, Beatson SA. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics. 2011;12(1):402. doi:10.1186/1471-2164-12-402
- Chang MX, Zhang JF, Sun YH, et al. Contribution of different mechanisms to ciprofloxacin resistance in Salmonella spp. Front Microbiol. 2021:12. doi:10.3389/fmicb.2021.663731
- Toleman MA, Bennett PM, Walsh TR. IS CR elements: novel gene-capturing systems of the 21st Century? Microbiol Mol Biol Rev. 2006;70(2):296–316. doi:10.1128/MMBR.00048-05
- Zhou Z, Alikhan NF, Mohamed K, et al. The EnteroBase user’s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny, and Escherichia core genomic diversity. Genome Res. 2020;30(1):138–152. doi:10.1101/gr.251678.119
- García-Soto S, Tomaso H, Linde J, Methner U. Epidemiological analysis of Salmonella enterica subsp. enterica serovar Dublin in German cattle herds using whole-genome sequencing. Microbiology Spectrum. 2021;9(2):e00332–21. doi:10.1128/Spectrum.00332-21
- Caliskan-Aydogan O, Alocilja EC. A review of carbapenem resistance in Enterobacterales and its detection techniques. Microorganisms. 2023;11(6):1491. doi:10.3390/microorganisms11061491
- Wu Y, Jiang T, Bao D, et al. Global population structure and genomic surveillance framework of carbapenem-resistant Salmonella enterica. Drug Resist Updates. 2023;68:100953. doi:10.1016/j.drup.2023.100953
- Wang Y, Liu Y, Lyu N, et al. The temporal dynamics of antimicrobial-resistant Salmonella enterica and predominant serovars in China. Natl Sci Rev. 2023;10(3):nwac269. doi:10.1093/nsr/nwac269
- Li Y, Li K, Peng K, Wang Z, Song H, Li R. Distribution, antimicrobial resistance and genomic characterization of Salmonella along the pork production chain in Jiangsu, China. LWT. 2022;163:113516. doi:10.1016/j.lwt.2022.113516
- Zhao Q, Berglund B, Zou H, et al. Dissemination of blaNDM-5 via IncX3 plasmids in carbapenem-resistant Enterobacteriaceae among humans and in the environment in an intensive vegetable cultivation area in eastern China. Environ Pollut. 2021;273:116370. doi:10.1016/j.envpol.2020.116370
- Tian D, Wang B, Zhang H, et al. Dissemination of the bla NDM-5 Gene via IncX3-Type Plasmid among Enterobacteriaceae in Children. mSphere. 2020;5(1):e00699–19. doi:10.1128/mSphere.00699-19
- Zeng S, Huang Y, Zhang X, Fu L, Sun Z, Li X. Molecular characterization of IncFII plasmid carrying bla NDM-5 in a Salmonella enterica serovar Typhimurium ST34 clinical isolate in China. mSphere. 2023;8(6):e00480–23. doi:10.1128/msphere.00480-23
- Johnson TJ, Thorsness JL, Anderson CP, et al. Horizontal gene transfer of a ColV plasmid has resulted in a dominant avian clonal type of Salmonella enterica serovar Kentucky. PLoS One. 2010;5(12):e15524. doi:10.1371/journal.pone.0015524
- Li IC, Wu HH, Chen ZW, Chou CH. Prevalence of IncFIB plasmids found among Salmonella enterica serovar schwarzengrund isolates from animal sources in Taiwan using whole-genome sequencing. Pathogens. 2021;10(8):1024. doi:10.3390/pathogens10081024
- Khajanchi BK, Hasan NA, Choi SY, et al. Comparative genomic analysis and characterization of incompatibility group FIB plasmid encoded virulence factors of Salmonella enterica isolated from food sources. BMC Genomics. 2017;18(1):1–14. doi:10.1186/s12864-017-3954-5
- Liu Z, Xiao X, Li Y, Liu Y, Li R, Wang Z. Emergence of IncX3 plasmid-harboring blaNDM–5 dominated by Escherichia coli ST48 in a goose farm in Jiangsu, China. Front Microbiol. 2019;10:2002. doi:10.3389/fmicb.2019.02002
- Li Y, Yang Y, Wang Y, Walsh TR, Wang S, Cai C. Molecular characterization of blaNDM-harboring plasmids reveal its rapid adaptation and evolution in the Enterobacteriaceae. One Health Adv. 2023;1(1):30. doi:10.1186/s44280-023-00033-9
- Corno G, Ghaly T, Sabatino R, et al. Class 1 integron and related antimicrobial resistance gene dynamics along a complex freshwater system affected by different anthropogenic pressures. Environ Pollut. 2023;316:120601. doi:10.1016/j.envpol.2022.120601
- Tavares RDS, Tacão M, Ramalheira E, Ferreira S, Henriques I. Report and Comparative Genomics of an NDM-5-Producing Escherichia coli in a Portuguese hospital: complex class 1 integrons as important players in blaNDM Spread. Microorganisms. 2022;10(11):2243. doi:10.3390/microorganisms10112243
- Chowdhury G, Ramamurthy T, Das B, et al. Characterization of NDM-5 carbapenemase-encoding gene (blaNDM-5) – positive multidrug resistant commensal Escherichia coli from diarrheal patients. IDR. 2022;15:3631–3642. doi:10.2147/IDR.S364526
- Fang L, Lin G, Li Y, et al. Genomic characterization of Salmonella enterica serovar Kentucky and London recovered from food and human salmonellosis in Zhejiang Province, China (2016–2021). Front Microbiol. 2022;13:961739. doi:10.3389/fmicb.2022.961739
- He Y, Wang J, Zhang R, et al. Epidemiology of foodborne diseases caused by Salmonella in Zhejiang Province, China, between 2010 and 2021. Front Public Health. 2023;11:1127925. doi:10.3389/fpubh.2023.1127925
- Authority EFS. The European Union One Health 2022 Zoonoses Report. EFSA J. 2023;21(12):1 doi:10.2903/j.efsa.2023.8442