
ABSTRACT
Fossil remains from the latest Messinian of Casino Basin (Tuscany, Italy) are known since the beginning of the nineteenth century. Among the others, the mammal assemblage includes Mesopithecus pentelicus, Tapirus arvernensis, Propotamochoerus provincialis and scarce and poorly preserved remains of a hippopotamid, consisting of a mandibular symphysis fragment, an apical fragment of a lower canine, some isolated lower incisors, a fragmented second upper premolar and a second lower molar. These specimens were initially referred as Hippopotamus hipponensis and later ascribed to the new species Hippopotamus pantanellii (recently reported as Hexaprotodon? pantanellii). However, this attribution has been disputed during the past years. The hippopotamid remains from the Casino Basin are revised here in order to clarify their systematic position and to infer paleobiogeographic and evolutionary patterns within the Mediterranean fossil record of Hippopotamidae. The morphology of the remains collected from the Casino Basin more closely resembles the African than the Asian hippopotamids’ lineage and therefore the Tuscan remains should be more properly referred as Archaeopotamus pantanellii. The latter species probably dispersed into Tuscany from the Iberian Peninsula where the presence of Archaeopotamus crusafonti is well documented.
Introduction
Hippopotamidae (Gray Citation1821) representatives appeared in the African and Arabic fossil record around 8 Ma (Boisserie et al. Citation2017b). The first and archaic Hippopotaminae include the poorly known genus Chororatherium, for now represented by a single described species, C. roobi from Chorora, Ethiopia (ca. 8 Ma) (Boisserie et al. Citation2017b). Starting from 7.5 Ma the ‘narrow-muzzle’ species belonging to the genus Archaeopotamus (Boisserie Citation2005; Boisserie et al. Citation2017a) started to be fairly represented in Africa and in the Arabian Peninsula. From 6 Ma onwards, the representatives of the genus Hexaprotodon Falconer and Cautley, Citation1836, which mainly comprises the Asian lineage of hippopotamids, also began to be common in mammal assemblages (Boisserie Citation2005). The poorly known genus Saotherium appeared in Chad (Central Africa) during the Early Pliocene (Boisserie et al. Citation2003; Boisserie Citation2005). In addition to the previous genera, Geraads et al. (Citation2021) ascribed some Plio-Pleistocene (2.9–2.4 Ma) remains from Mille‐Logya in the Lower Awash Valley, Ethiopia, to ‘Trilobophorus’ but its validity needs more evidence. Further studies also need to be carried out on Pliocene materials from Kenya, Hadar (ca. 3 Ma), Kanapoi (ca. 4 Ma), lower parts of Koobi Fora Formation at Allia Bay (4.2–4.1 Ma), Nachukui Formation (Apak Member, more than 4.3 Ma), Omo Group formations (3.4–2.9 Ma) and Kantis (3.4–3.5 Ma), where most of the remains are currently ascribed as aff. Hippopotamus (Gèze Citation1985; Harris 1991; McDougall and Feibel Citation1999; Harris et al. Citation2003; Weston Citation2003; McDougall and Brown Citation2008; Mbua et al. Citation2016). The North African peri-Mediterranean area during that time frame was marked by several hippopotamids occurrences. Hexaprotodon? sahabiensis was described from As Sahabi, Libya, Late Miocene (Gaziry Citation1987; Pavlakis Citation2008), Hex.? hipponensis from Pont-de-Duvivier, Algeria, earliest Pliocene (Gaudry Citation1876) and Hex.? protamphibius andrewsi from Wadi Natrun, Egypt, Late Miocene-Early Pliocene (Andrews Citation1902; Stromer Citation1914; Arambourg Citation1947). Hexaprotodon? sahabiensis material is quite abundant, mainly represented by isolated teeth and a fragment of a mandible with right canine, right incisors and the left first incisor (Gaziry Citation1987; Pavlakis Citation2008). Hexaprotodon? sahabiensis is quite peculiar and it probably represents an evolutionary branch with both archaic and advanced characteristics (Gaziry Citation1987). Hexaprotodon? hipponensis was established on scarce and fragmentary material, including some isolated incisors, a premolar and a canine fragment (Gaudry Citation1876). Hexaprotodon? protamphibius andrewsi is represented by numerous cranial, mainly isolated teeth, and post-cranial remains (Andrews Citation1902; Stromer Citation1914; Arambourg Citation1947). Lately, Hex.? protamphibius andrewsi was reassigned to Archaeopotamus andrewsi, as an extensive revision based on all the abundant material collected from Wadi Natrun (Egypt) revealed a closer relationship with Archaeopotamus rather than with Hexaprotodon (Pickford et al. Citation2022).
The first dispersal wave of Hippopotamidae outside Africa towards Europe took place around 6 Ma (Boisserie Citation2007). In the European area, hippopotamids remains were collected from several Late Miocene deposits of Spain (Aguirre Citation1963; Crusafont et al. Citation1964; Aguirre et al. Citation1973; Morales Citation1984; Lacomba et al. Citation1986; Alcalá Citation1994; Alcalá and Montoya Citation1998; Morales et al. Citation2011), from an Early Pliocene locality of France (Faure and Méon Citation1984) and from latest Miocene sites of Italy (Pantanelli Citation1879; Seguenza Citation1902, Citation1907; Hooijer Citation1946; Martino et al. Citation2021). Hippopotamid occurrence in the Late Miocene deposits of Greece, at Panaghía sta Éria (Euboea Island), should be considered doubtful (Athanassiou Citation2022). The rich hippopotamid material, represented by cranial and post-cranial remains, collected from Spain (Las Casiones, Venta del Moro, La Portera, Arenas del Rey, El Arquillo I) is ascribed to Hex.? crusafonti (=Hex. primaevus) (Aguirre Citation1963; Crusafont et al. Citation1964; Aguirre et al. Citation1973; Morales Citation1984; Lacomba et al. Citation1986; Alcalá Citation1994; Alcalá and Montoya Citation1998; Morales et al. Citation2011). However, in a recent revision, Martino et al. (Citation2021) suggested that this Late Miocene species should be referred to the genus Archaeopotamus, based on the peculiar mandibular characters shared by the material from La Portera (Lacomba et al. Citation1986). The remains ascribed to Hex.? crusafonti should therefore be more properly indicated as A. crusafonti. The latter species is also reported from La Mosson (France), dated to the earliest Pliocene (Faure and Méon Citation1984).
In Italy, between the end of the nineteenth century and the beginning of the twentieth one, Late Miocene hippopotamid remains were found in four localities (Pantanelli Citation1879; Seguenza Citation1902, Citation1907). Seguenza (Citation1902, Citation1907) described a considerable number of fossils collected from Gravitelli (Messina, Sicily, southern Italy) and some isolated remains unearthed from two different localities near Gravitelli, San Pier Niceto and Scirpi (Messina, Sicily, southern Italy). Unfortunately, all the fossil records collected went lost in 1908 due to a catastrophic earthquake. The remains collected from Sicily are currently dubiously assigned to Hex.? siculus (Seguenza Citation1902, Citation1907; Hooijer Citation1946; Boisserie Citation2005; Martino et al. Citation2021). Some years before Seguenza (Citation1902, Citation1907), Pantanelli (Citation1879) reported some fragmentary Late Miocene hippopotamid material from a Tuscan locality, the Casino Basin (Siena). These remains were originally assigned by Pantanelli (Citation1879) to Hippopotamus hipponensis Gaudry, Citation1876 (now Hex.? hipponensis following Boisserie Citation2005). Hereafter Joleaud (Citation1920) revised the material described from the Casino Basin and agreed with Stehlin (Citation1899-Citation1900), Forsyth Mayor (Citation1875) and Andrews (Citation1902) in considering ‘la forme de Casino diffère de celle de Duvivier et confine plus directement aux Suilliens par le dessin de la surface d’usure des molaires qui ne présentent pas encore la tréfle caractéristique des Hippopotames pliocène, quaternaires et actuels’ (the Casino specimens differ from the material from Pont-de-Duvivier by the design of the molars, which do not yet display the characteristic trefoil wear pattern of the Pliocene, Quaternary and modern hippopotamuses). For the abovementioned reasons, Joleaud (Citation1920) rejected the attribution to Hip. hipponensis (now Hex.? hipponensis) and assigned the poorly represented material from the Casino Basin to a new species, Hippopotamus pantanellii Joleaud (Citation1920). An extensive revision by Boisserie (Citation2005) doubtfully attributed, on the base of the tables in Pantanelli (Citation1879), the Tuscan remains to the genus Hexaprotodon, rejecting the attribution to the more evolved Hippopotamus.
The original hippopotamid remains collected from the Casino Basin are here revised in detail and figured for the first time in order to clarify and update their systematic position and their paleobiogeographic implications. Despite the fragmentary nature of the remains collected from the Casino Basin, a detailed revision is important to shed light on the scant Late Miocene hippopotamid material collected from the peri-Mediterranean area. Hippopotamids, thanks to their peculiar semi-aquatic lifestyle, represent a unicum among large mammals and therefore they can contribute for a better understanding of the great changes that affected the peri-Mediterranean area during the Late Miocene.
Geological setting
Fossil presence in the Casino Basin, Tuscany, Central Italy (Casino Basin location is indicated with the black star in ) was firstly reported by Capellini (Citation1872). Casino Basin’s area (around 40 km2) is mainly characterised by two different sedimentary sequences. The geological setting of the Casino Basin was firstly defined as a combination of an Upper Lacustrine Cycle and a Lower Lacustrine Cycle (Lazzarotto and Sandrelli Citation1977). Bossio et al. (Citation2002) indicated the Upper Lacustrine Cycle as ‘Argille del Casino’ (Casino clays). Later, Abbazzi et al. (Citation2008) recognised a unit lithologically and palaeontologically coinciding with Casino clays along the Borro Strolla Creek. The UM2 lithofacies, outcropping in Piaggiole A quarry of the Borro Strolla synthem, deposited simultaneously to similar sedimentation in the second Casino’s Lake (Abbazzi et al. Citation2008). This unit, as the Casino clays, can be ascribed to the latest Messinian, ‘Lago-Mare’ bio-facies.
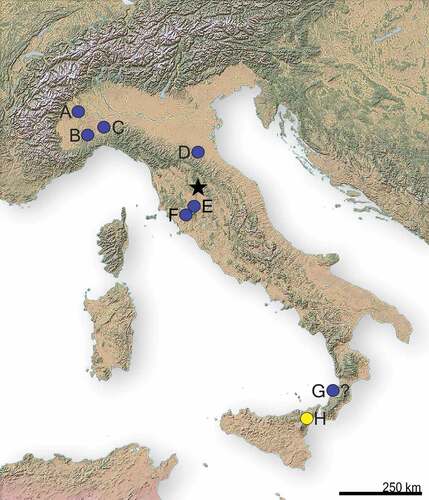
Figure 1. Casino Basin locality indicated with the black star. A. Moncucco Torinese, B. Verduno, C. Ciabot Cagna, D. Monticino Quarry, E. Velona, F. Baccinello V3, G. Cessaniti, H. Sicilian localities (Gravitelli, San Pier Niceto, Scirpi). Borro Strolla locality coincides with Casino Basin one. Yellow = hippopotamids presence, blue = hippopotamids absence.
The faunal list of the second Casino Lake includes Dipoides problematicus, Eucyon sp., Thalassictis cf. hipparionum, Tapirus arvernensis, Propotamochoerus provincialis, Parabos sp., Paracervulus cf. australis, Hippopotamidae indet., Mesopithecus pentelicus and Hippotherium cf. malpassi (Rook Citation1992; Rook et al. Citation1999; Rook and Bernor Citation2013; Cirilli et al. Citation2020; Martino et al. Citation2020; Iannucci et al. Citation2021; Pandolfi et al. Citation2021a, Citation2021b). Some remains display bitemarks on posterior bones, and carnivores may have therefore played a role in their accumulation (Gallai Citation2005). The mammals’ assemblage of the Casino Basin can be ascribed to the upper part of the Mammal Neogene Zone 13.
Material and methods
The morphological terminology for the teeth followed Thenius (Citation1989) and Boisserie et al. (Citation2010). The revised remains were morphologically and morphometrically compared with Late Miocene and Early Pliocene hippopotamids from direct observations and published data. The species analysed from bibliography are Archaeopotamus harvardi (Coryndon Citation1977), Archaeopotamus qeshta (Boisserie et al. Citation2017a), Archaeopotamus lothagamensis (Weston Citation2000), Archaeopotamus crusafonti (Aguirre Citation1963), Archaeopotamus andrewsi (Arambourg Citation1947), Hexaprotodon bruneti (Boisserie and White, Citation2004), Hexaprotodon garyam Boisserie, Likius, Vignaud, & Brunet, Citation2005, Hexaprotodon sivalensis Falconer & Cautley, Citation1836, Hexaprotodon? hipponensis (Gaudry, Citation1876), Hexaprotodon? sahabiensis (Gaziry Citation1987), Hexaprotodon? siculus (Hooijer Citation1946) and Saotherium mingoz Boisserie, Citation2005. R.M. studied A. crusafonti remains stored in the Museo Nacional de Ciencias Naturales of Madrid (Spain), collected from Arenas del Rey, El Arquillo and La Portera. All specimens are reported in Aguirre (Citation1963), Lacomba et al. (Citation1986) and Alcalá and Montoya (Citation1998). The specimens from the Casino basin were also compared with the remains of Hippopotamus amphibius Linnaeus, Citation1758 and Choeropsis liberiensis (Morton Citation1844).
Historical framework
Fossiliferous remains from the Casino Basin were firstly reported by Capellini (Citation1872), Rütimeyer (Citation1876) and Major (Citation1874, Citation1877). A more detailed study regarding the Casino Basin was tackled by Pantanelli (Citation1879, Citation1886). Pantanelli (Citation1879) briefly described and depicted the faunistic remains from the Casino Basin and ascribed the hippopotamid remains to Hip. hipponensis (now Hex.? hipponensis), previously described by Gaudry (Citation1876) in Algeria. However, the morphology of the material figured by Pantanelli (Citation1879) was highly simplified. An attempt to revise the hippopotamid remains from the Casino Basin was made by Joleaud (Citation1920). However, the author never revised the original specimens from the Casino Basin, but only the description and tables depicted in Pantanelli (Citation1879). Joleaud (Citation1920) erected the new species, Hip. pantanellii, in honour of Dante Pantanelli, who firstly described these remains. However, Joleaud (Citation1920) never described the diagnostic characters of Hip. pantanellii, which are still currently undefined. All the remains collected from the Casino Basin are now stored in the Museo di Storia Naturale dell’Accademia dei Fisiocritici, Siena, Italy, with the exception of the second lower molar that is currently in the Museo di Storia Naturale e del Territorio, Certosa di Calci, Pisa, Italy.
Institutional Abbreviations
MSNAF, Museo di Storia Naturale dell’Accademia dei Fisiocritici, Siena, Italy; MSNCC, Museo di Storia Naturale e del Territorio, Certosa di Calci, Pisa, Italy; MNCN, Museo Nacional de Ciencias Naturales, Madrid, Spain.
Other Abbreviations
C/c, canines; I/i, incisors; P/p, premolars; M/m, molars; L, greatest length; W, width; MD, mesiodistal diameter; LL: labiolingual diameter; MN, Mammal Neogene Zone.
Systematic Palaeontology
Class Mammalia Linnaeus Citation1758
Cetartiodactyla Montgelard et al. Citation1997
Superfamily Hippopotamoidea Gray Citation1821 (sensu Gentry and Hooker Citation1988)
Family Hippopotamidae Gray Citation1821
Subfamily Hippopotaminae Gray Citation1821
Genus Archaeopotamus Boisserie, Citation2005
Type species. Archaeopotamus lothagamensis (Weston Citation2000)
Included species
Archaeopotamus harvardi (Coryndon Citation1977); A. qeshta (Gentry Citation1999; Boisserie et al. Citation2017a; Boisserie and Bibi Citation2022); A. crusafonti (Aguirre Citation1963; Crusafont et al. Citation1964; Aguirre et al. Citation1973; Morales Citation1984; Lacomba et al. Citation1986; Alcalá Citation1994; Alcalá and Montoya Citation1998; Morales et al. Citation2011); A. andrewsi (Andrews Citation1902; Stromer Citation1914; Pickford et al. Citation2022); A. aff. harvardi from Rawi (see Boisserie Citation2005).
Archaeopotamus pantanellii (Joleaud, Citation1920) nov. comb.
()
Figure 2. A. Mandible fragment with right i1, left i1, i2, i3 (MSNAF2821). B-E, right lower canine (MNSAF2820). B, lateral view; C, occlusal view; D, mesial view; E, cross section. F-I, lower incisors (MSNAF2829). F, i2-i3; G, i1-i2; H, i1-i2; I, lower undeterminable incisors fragments. J-L, right P2 (MSNAF2828). J, lingual view; K, labial view; L, occlusal view. M, lower premolar fragment (p1?) (MSNAF2828). N-R, left m2 (MSNCCI10501). N, labial view; O, distal view; P, occlusal view; Q, mesial view; R, lingual view. Scale bars 2 cm.

Figure 3. Original table of Casino Basin remains (Pantanelli Citation1879), Tab. IV, Figures 1–7. 1–2, MNSAF2820. 3, MNSAF2828. 4, MSNCCI10501. 5, MNSAF2820. 6–7, MNSAF2829.
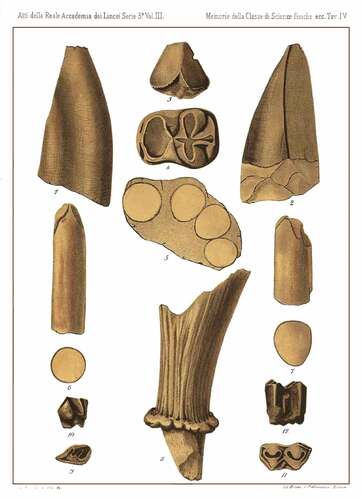
1879: Hippopotamus hipponensis in Pantanelli Citation1879, p. 318, plate IV, 1–7.
1920: Hippopotamus pantanellii in Joleaud Citation1920, p. 18.
2005: Hexaprotodon? pantanellii in Boisserie Citation2005, p. 20.
Material
Holotype, MSNAF2821, fragmented mandible with right i1, left i1, i2, i3 in cross-section. The rest of the remains from the type locality are defined as paratypes.
Other referred specimens
MSNAF2829, isolated lower incisors; MSNAF2820, apical fragment of a right c; MSNAF2828, a right P2 and severely fragmented tooth (probably a p1); MSNCCI10501, left m2; MSNAF2862, fragmented mandible with undeterminable teeth in cross-section. All remains probably belong to at least two different individuals, a sub-adult (p2 and c) and an adult (mandible and m2).
Derivation of name
Archaeopotamus pantanellii, in honour of Dante Pantanelli, the first scholar who described the hippopotamid from the Casino Basin.
Stratigraphical range
Late Miocene (MN 13).
Type locality
Upper lacustrine level of Casino Lignite mine (also known as Casino clays or second Casino’s Lake), near Siena, Tuscany, Italy (coordinates 43.3° N, 11.3° E).
Emended diagnosis
Archaeopotamus pantanellii is a medium-small European hexaprotodont species clearly distinguished from other hippopotamids by the following combination of characters: incisors arrangement i1 > i2 ≥ i3 with rounded-oblate cross-section; c with a D-shaped cross-section, a groove on the mesial side, finely crenulated enamel, and slightly visible transversal lines; P2 triangular shaped without accessory cuspids; m2 with four main cuspids, well-developed cingulid on lingual, mesial and distal sides, crenulated enamel and feebly developed trefoil wear pattern.
Differential diagnosis
Archaeopotamus pantanellii diverges from A. harvardi in having transversal fine lines on the c; the former species is also characterised by smaller dimensions. Archaeopotamus pantanellii differs from A. lothagamensis in showing a groove on the c mesial side and larger dimensions and can be distinguished from A. qeshta for the i3> i2≥ i1 arrangement. Archaeopotamus pantanellii, differently from A. andrewsi, shows transversal lines on the c enamel and a m2 with acingulid more crenulated. Archaeopotamus pantanellii differs from A. crusafonti in being hexaprotodont and showing overall larger dimensions, the Tuscan species also differs from Hex.? siculus in having a single groove on the mesial side of the c and straight incisors without grooves. Archaeopotamus pantanellii differently from Hex.? sahabiensis displays m2 with four main cuspids instead of five. Archaeopotamus pantanellii is recognisable from Hex.? hipponensis for straight incisors without peculiar wear surface.
General description
The mandible (MSNAF2821) is highly damaged and displays four teeth in cross-section that could be right i1, left i1, i2, i3 or right c, i3, i2 and i1 (). The mandibular fragment is probably a part of the mandibular symphysis, close to the teeth emergence. Among the two options, probably the first one is the more conservative. The putative right i1 does not show a different internal structure or visible enamel, and the cross-section is likewise similar to the other teeth. In cross-section the putative i3 is the smallest incisor, and the i2 displays slightly smaller dimensions to i1, whilst the i1 is the largest one. A small diastema is present between the two i1s. The i2 and the i3 have rounded cross-section, while the i1s are more elliptic-shaped. The diastema between the right i1 and the left i1 is well visible, while it is absent between i1 and i2 and i2 and i3. In the light of that, the Casino Basin hippopotamid was thus characterised by a hexaprotodont condition. A lignite vein well visible along the mandibular fragment could attest the possible action of intense taphonomic activities that could have altered the original morphology of the remains. The action of post-depositional processes could explain the partially folded aspect of the incisors in the mandibular fragment.
The right c is severely fragmented and only represented by the apical part MSNAF2820 (). The mesial side () is better preserved than the lateral one (). On the former side is visible a longitudinal groove that runs on the upper-central part of the canine, no similar structure is visible on the lateral side. The enamel is thin, finely crenulated with peculiar slightly visible transversal lines. The apical part in the occlusal view () displays a deflection towards the mesial side. The cross-section (), although if incomplete because it includes the wear surface, shows a cross-section that is mostly D-shaped, and it presents a small groove on the mesial side that contributes to a bean-shaped aspect of the cross-section.
The isolated lower incisors (), three well preserved () and two undetermined fragments (), are mainly cylindrical with a round or mostly rounded cross-section (MSNAF2829). All the incisors lack the distal part. The enamel is just slightly visible on some specimens (). All remains show an apical bevelled wear facet. One incisor () is long and slender, while the other two are smaller and with a stockier aspect (). The incisor depicted in has a cross-section more elliptic, with a mesial-lateral diameter greater than the ventral-dorsal one.
The right P2 MSNAF2828 () is broken along the longitudinal plane, distal and mesial parts are partially missing, roots are almost entirely damaged and therefore not investigable. The P2 is mainly triangular and does not show accessory cusps, the enamel is finely crenulated. The cingulum is pustulated on the lingual side (). On the labial side (), the cingulum looks less developed than on the lingual side. In occlusal view (), the main cusp of the P2 curves lingually towards the apex. The P2 is just slightly worn. On the lower part of the lingual side are visible several transversal thin bands. The same bands are also visible on the other fragmented premolar (probably a lower p1, ). These bands are probably growth lines (perikymata).
The left m2 is transversally damaged MSNCCI10501 (). The enamel is rough, and accessory cuspids are absent. The hypoconid in occlusal view is the largest cuspid, and it is linked with the mesial cuspids (metaconid and protoconid) as well as the cingulid distally (). The metaconulid is reduced, mostly ovoid shaped. The cingulid is well developed on the labial part (), particularly on the protoconid. The cingulid is almost visible on the lingual part, poorly developed on the metaconid and absent on the entoconid (). The cingulid is particularly high and slightly pustulated on the distal part of the tooth (), while less developed on the mesial part (). On the medial labial side is present a mesostylid. The m2 looks fairly worn in occlusal view () in particular, on the anterior cusps. The distal part is better preserved, and the trefoil wear pattern typical of the Hippopotamidae family is almost completely developed. The roots are nearly entirely damaged.
The last remain is probably another mandible part; only two cross-sections of indeterminable incisors are visible that are neither diagnostic nor significant for an anatomical attribution (MSNAF2862).
Morphological and morphometrical comparison
Morphological comparison
A comparison with the hippopotamids unearthed from coeval localities is not easy due to the fragmentary and scant nature of the material collected from the Casino Basin. The morphology of the mandible fragment was highly simplified in Pantanelli (Citation1879) (). The cross-sections of the teeth were depicted as perfectly sub-rounded, although the real morphology is more complex. The largest incisor in the mandibular fragment is the i1, while the i2 and the i3 appear dimensionally similar, with the i2 slightly larger than the i3. This incisors arrangement (i1 > i2 ≥ i3) is quite common in hippopotamids. Archaeopotamus harvardi displays the i1 bigger than i2 and i3, while in A. qeshta i2 is the largest incisor (Weston Citation2000, Citation2003; Boisserie et al. Citation2017a). Boisserie et al. (Citation2017a) reported an i3> i2> i1 arrangement for A. lothagamensis, while, conversely, Weston (Citation2000) described these incisors as equal-sized. However, the measurements reported in Boisserie et al. (Citation2017a) supported the i3> i2> i1 condition. Archaeopotamus crusafonti displays only four incisors in the mandible, with the i1 slightly larger than the i2 (Lacomba et al. Citation1986; Martino et al. Citation2021). Hex. sivalensis usually has the i2 smaller than the i1, while the i3 in some specimens can be bigger than the i1 (Boisserie et al. Citation2005b). For de Visser (Citation2008), in Asian species belonging to Hexaprotodon, the relative size of the lower incisors may be variable. Hexaprotodon garyam shows the i1 and the i3 equal in size or the i1 slightly larger than the i3, while the i2 is always the smallest one (Boisserie et al. Citation2005b). Hex. bruneti is characterised by a particularly enlarged i3 and by i2 and i1 similar in size (Boisserie and White Citation2004). The relative dimension of the six incisors in the mandible of Hex.? siculus is unknown (Seguenza Citation1902, Citation1907; Martino et al. Citation2021). In Hex.? hipponensis the i1 is reported as the largest incisor (Gaudry Citation1876). Nevertheless, a mandible of Hex.? hipponensis with incisors in situ has never been recovered, and therefore this assertion is not necessarily correct. Hexaprotodon? sahabiensis has the i2 smaller than the i1 and the i3, whilst the i1 and the i3 are almost sub-equal in size (Pavlakis Citation2008). Soatherium mingoz shows similar dimensions for the i1 and the i3, while the i2 is always the smallest incisor (Boisserie et al. Citation2003). Hippopotamus amphibius usually displays a tetraprotodont condition, with the i1 bigger than the i2. Nevertheless, Falconer (Citation1868, p. 406 footnote), Gaudry (Citation1876) and Stuenes (Citation1989) described a supernumerary incisors’ anomaly in some H. amphibius specimens affecting one side of the lower jaw in adult individuals. Choeropsis liberiensis is characterised by a diprotodont condition, and the mandible has therefore only two first incisors. The morphology of the lower incisors is likewise poorly informative. Weston (Citation2003) stated that in A. harvardi the morphology is variable, some isolated incisors collected from Lothagam are curved, with longitudinal lateral grooves (LT-26207), while others have a wear facet highly irregular. Archaeopotamus crusafonti i2 (MNCN72803) reported in Alcalà and Montoya (Citation1998), is mainly cylindrical with a flattened mesial side, almost straight and with a cross-section that shows a feeble groove on the lateral side, similarly to the specimen depicted in . The fragmented left hemimandible (MNCN62837) displays the cross-section of the alleged i1 slightly elliptical, a general cylindrical aspect, and two poorly developed grooves on the labial and mesial sides, respectively. Archaeopotamus andrewsi had mainly cylindrical incisors with a smooth or slightly grooved enamel (Stromer Citation1914; Pickford et al. Citation2022). Nevertheless, one incisor (p. 54, fig. 85, D1-D2) depicted by Pickford et al. (Citation2022) displays the ‘tracked margin’ of the wear surface typical of Hex.? hipponensis (Gaudry Citation1876). In Hex. bruneti the i1 has a circular cross-section, whereas the i2 may be mesial-laterally compressed. The i3, the largest incisor, can exhibit both a circular cross-section and a mesial-lateral compressed one (Boisserie and White Citation2004). Hexaprotodon? sahabiensis mandible described in Pavlakis (Citation2008) displays large incisors with a simple morphology, cylindrical and straight. Pavlakis (Citation2008) did not describe the enamel surface of the lower incisors in Hex.? sahabiensis. The incisors of Hex.? siculus depicted by Seguenza (Citation1907, Tab. VII figs. 13–16) probably belonged to a single individual, since the author referred to a mandible, destroyed during the excavation, with several teeth in situ. Some of these incisors were slightly apically curved (figs 14 and 16), others exhibited a well-visible groove (figs. 14 and 15), while one (fig. 13) was mainly cylindrical. The morphology of the incisors collected from the Casino Basin is probably one of the simplest among all the Late Miocene hippopotamids.
The c from the Casino Basin displays a longitudinal groove on the mesial side and an enamel finely crenulated, similarly to A. harvardi (Weston Citation2003). In A. harvardi, the cross-section of the c is slightly bean-shaped and, despite the cross-section of the c from the Casino Basin is not fully investigable, the D-section resembles the one displayed by A. harvardi (Weston Citation2003, p. 456, fig. 10.16). In A. harvardi collected from Tanzania the c cross-section is more D-shaped than bean-shaped (Harrison Citation1997). Archaeopotamus lothagamensis has a c compressed from side to side with an enamel finely striated and a flattened mesial border (Weston Citation2000). No grooves are reported by Weston (Citation2000) on the severely fragmented c described from Lothagam (Kenya). The cross-section of the c of A. lothagamensis is mainly D-shaped (Weston Citation2000). The c from the Casino Basin is similar, but less transversally compressed. The fine striated enamel similarly characterised A. qeshta, which displays a bean-shaped cross-section and a flat-to-concave mesial side (Boisserie et al. Citation2017a). Archaeopotamus crusafonti c (MNCN72765) displays thin finely crenulated enamel, a groove on the mesial side that is less marked than in H. amphibius and an additional groove barely developed more ventrally (pers. observ. R.M.). The cross-section of the c is visible on the mandibular symphysis collected from La Portera (MNCN62837), it is bean-shaped with a groove on the mesial side and a more rounded aspect on the lateral side. Archaeopotamus andrewsi c has a flat mesial side and a more rounded lateral one, the enamel is finely striated, and a well-visible groove runs along the mesial side (Pickford et al. Citation2022). The groove on the c of A. andrewsi (Pickford et al. Citation2022, p. 55, fig. 86 A and p. 60, fig. 97) seems narrower and deeper than in the Casino Basin specimen. A small groove on the lateral side of the A. andrewsi c (Pickford et al. Citation2022, p. 55, fig. 86 B) is well visible, but the latter is placed more ventrally than in the Casino Basin specimen. The cross-section of the c in A. andrewsi is mainly D-shaped, as in most Archaeopotamus specimens (Pickford et al. Citation2022). Regarding Hexaprotodon genus, Hex. garyam has a finely wrinkled enamel, while Hex. sivalensis can show fine, coarse or smooth enamel (Boisserie et al. Citation2005b; de Visser Citation2008). Iqbal et al. (Citation2019) described lower canines relatively well striated and with shallow grooves. de Visser (Citation2008) reported the presence of a groove on the lateral side on several specimens of Hexaprotodon, while the mesial side may be grooved or smooth. Nevertheless, on many canines it was not possible to investigate grooves’ occurrence (de Visser Citation2008). In Hex. sivalensis the c has a peculiar ‘pear-shaped’ cross-section (Falconer and Cautley Citation1868; de Visser Citation2008). The c cross-section in the Casino Basin is markedly different from that the one of Hex. sivalensis. The canine of Hex. bruneti is characterised by a gently wrinkled enamel (Boisserie and White Citation2004). Hexaprotodon? siculus c displays well-visible striae on the enamel and two grooves on the supposedly mesial side, the only one figured by Seguenza (Citation1907). However, Seguenza (Citation1907) described an additional groove on the lateral side, missing in the Casino Basin specimen. Hexaprotodon? sahabiensis is characterised by a significantly large c, bigger than A. harvardi and Hex. sivalensis, with finely crenulated or smooth enamel (Gaziry Citation1987). The mesial side of the c of Hex.? sahabiensis is flat, while the lateral one is more rounded, the c cross-section is defined as ‘kidney-shaped’, and it is peculiar among hippopotamine (Gaziry Citation1987; Pavlakis Citation2008). The enamel is finely crenulated or almost smooth, no lateral grooves were mentioned (Gaziry Citation1987; Pavlakis Citation2008). Gaudry (Citation1876) reported fine striations and no strong grooves on the c lateral side of Hex.? hipponensis (Gaudry Citation1876, p. 501, fig. 5). Arambourg (Citation1944) figured the c mesial side of Hex.? hipponensis, the latter is characterised by fine striae, while distinctive grooves are not detectable. The enamel of the c of S. mingoz can be finely ridged or smooth, and grooves on lateral and mesial sides were not reported by Boisserie et al. (Citation2003). The enamel of the c of C. liberiensis is finely striated, almost smooth. The c of the Hip. amphibius is longitudinally well striated, with grooves on both mesial and lateral sides. The groove on the mesial side is usually more developed than the one on the lateral one. The groove on the lateral side of the c is more or less developed in several hippopotamid species and usually well expressed in later ontogenetic stages (Boisserie Citation2005).
The P2 (MSNAF2828) is not well preserved. The outline is triangular, with no visible accessory cuspids. However, the mesial and distal sides are partially missing, so possible small accessory cusps could have been lost. P2s of A. harvardi are usually characterised by a single cusp curved lingually towards the apex and by a well-developed cingulum, in particular on the lingual side. In addition, pustulated enamel can be seen running mesio-lingually and disto-labially (Weston Citation2003). These characters are shared also by the hippopotamid from the Casino Basin. Currently, no P2s of A. lothagamensis, A. crusafonti and A. qeshta are known (Lacomba et al. Citation1986; Weston Citation2000; Boisserie et al. Citation2017a). Regarding Hex. garyam, Boisserie et al. (Citation2005b) reported a P2 with a main triangular cusp, a strong mesial and distal cingulum as well as a mesial crest lingually curved. de Visser (Citation2008) highlighted great variability in upper premolars of Hexaprotodon, P2 usually displays one main cusp and in most of the teeth is also present a posterior platform, covered by crenulated ridges or cusplets, and the latter character is absent in the specimen collected from the Casino Basin. Hex. bruneti P2 is bad preserved, and therefore its morphology is unknown (Boisserie and White Citation2004). Soatherium mingoz has a triangular P2 in lateral view (Boisserie et al. Citation2003). Hex.? siculus is characterised by a simple-built P2, with a gently wrinkled enamel (Martino et al. Citation2021). The P2s of Hex.? sahabiensis and Hex.? hipponensis are both unknown (Gaziry Citation1987; Pavlakis Citation2008). The P2 of H. amphibius is mainly triangular and monocusp P2 in C. liberiensis appears mainly triangular, with a possible distal accessory cusp, wrinkled enamel and pustulated cingulum.
The m2 from the Casino Basin (MSNCCI10501) was rediscovered recently in the MSNCC. The morphology of the m2 depicted in the original table (Pantanelli Citation1879, Tab. IV, fig. 4), is highly simplified (). Hooijer (Citation1946) argued that the m2 collected from the Casino Basin shows the hypoconid more simply built than the entoconid, differently from Hippopotamus and Asian Hexaprotodon. In Hippopotamidae, the entoconid is usually characterised by a comma-shaped wear surface, differently from the hypoconid that is usually trifoliated and therefore more complex (Mazza Citation1995). The lower molars of A. harvardi are characterised by a great variation. In general, these molars possess four low cusps with a prominent cingulid that forms shelves mesially and distally. In A. harvardi the entoconid is usually comma-shaped (Weston Citation2003). In A. lothagamensis a medial stylid is present on the m2 that also possesses a well-developed distal cingular shelf, the entoconid is comma shaped (Weston Citation2000, Citation2003). Weston (Citation2000) described a hemi-mandible from Lothagam ascribed to A. cf. lothagamensis (KNM-LT 23871). The m2 from this mandible is similar in occlusal view to the one from the Casino Basin in displaying a large hypoconid and a more ovoid-shaped entoconid. Archaeopotamus qeshta displays a m2 morphologically similar to the one from the Casino Basin (Boisserie et al. Citation2017a, p. 5, fig. 3B). The m2 of A. qeshta is slightly more worn than the one from Tuscany, but a large hypoconid and a smaller entoconid are well distinguishable. This m2 probably possesses a mesostylid on the labial part. Boisserie et al. (Citation2017a) provided just the occlusal view of the m2 of A. qeshta, therefore it is not possible to investigate the labial and lingual sides. An unworn m2 of A. crusafonti is described in Alcalá and Montoya (Citation1998). This tooth (MNCN72805) displays a well-developed cingulid, in particular on the mesial and distal parts. The hypoconid is way larger than the entoconid, the cingulid is high and crenulated distally and mesially, the cingulid is not high on both sides and it is also well visible on a mesostylid. In A. andrewsi the second lower molars depicted in Pickford et al. (Citation2022) are almost unworn, and thus comparison with the Tuscan specimen is difficult. However, the labial side (Pickford et al. Citation2022, p. 53, fig. 107, A3) displays a cingulid that looks less developed and less crenulated than in the m2 from the Casino, and in occlusal view (Pickford et al. Citation2022, p. 53, fig. 107, B1) the hypoconid looks noteworthy larger than the entoconid. On the m2s of Hex. sivalensis are visible an anterior lobe, some posterior cuspids (heptaconid, pentaconid, hexaconid), and most of them do not display cingulid (de Visser Citation2008). An accessory cuspid and the absence of cingulid on the m2 of Hex. sivalensis is also reported by Iqbal et al. (Citation2019). These characters of Hex. sivalensis are not shared with the hippopotamus collected from the Casino Basin. The m2 of Hex. garyam is characterised by a trefoil wear pattern not fully developed and a cingulid thicker mesially and distally but attenuated laterally. In Hex. bruneti, the cingulid is well developed and the entoconid has a mesial lobe (Boisserie and White Citation2004). The m2 of Hex.? siculus was only figured by Seguenza (Citation1907) in the lingual and labial views. The m2 of the Sicilian species displayed a cingulid better developed on the lingual side rather than on the labial one. The cingulid is quite crenulated and it is high mesially and distally (Seguenza Citation1907). Hexaprotodon? sahabiensis is characterised by five well-developed cuspids on the m2 (Gaziry Citation1987). In S. mingoz, the m2 is low-crowned and the cingulid is missing lingually and labially, low and thick mesially and distally (Boisserie et al. Citation2003). Choeropsis liberiensis displays an m2 with four cuspids, trefoil wear pattern, high cingulid on the anterior cusps of the m2, while less developed in the posterior ones. In H. amphibius the m2 is high crowned, with a trefoil wear pattern perfectly developed, high cingulid, four main cuspids and sometimes by an accessory distal cuspid. The main characteristics of Late Miocene and Early Pliocene species are listed in .
Figure 4. Paleobiogeographical reconstruction of Late Miocene-Early Pliocene localities with hippopotamids from the peri-Mediterranean area, modified from Popov et al. (Citation2004). For further information regarding colours and symbology check Map. 9 (latest Miocene), Contents Annexe in Popov et al. (Citation2004). A, Archaeopotamus pantanellii, Casino Basin, Tuscany, Italy; B, Archaeopotamus crusafonti, La Mosson, Montpellier, France; C, Archaeopotamus crusafonti, Las Casiones, Teruel, Spain; D, Archaeopotamus crusafonti, El Arquillo, Teruel, Spain; E, Archaeopotamus crusafonti, Venta del Moro and La Portera, Valencia, Spain; F, Archaeopotamus crusafonti, Arenas del Rey, Granada, Spain; G, Hexaprotodon? hipponensis, Pont-de-Duvivier, Algeria; H, Hexaprotodon? siculus, Gravitelli, Scirpi, San Pier Niceto, Sicily, Italy; I, Hexaprotodon? sahabiensis, As Sahabi, Libya; J, Archaeopotamus andrewsi, Wadi Natrun, Egypt.
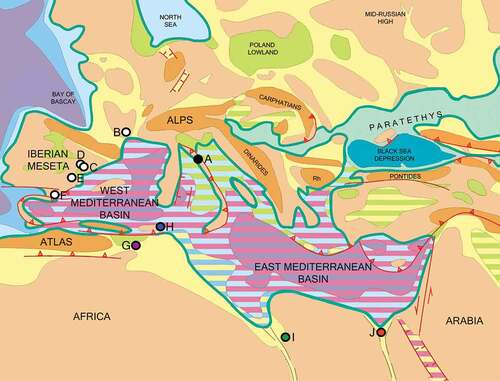
Table 1. Characters shared by A. pantanellii compared to Late Miocene and Early Pliocene hippopotamids. Archaeopotamus harvardi (Harrison Citation1997; Weston Citation2003), A. lothagamensis (Weston Citation2000, Citation2003), A. qeshta (Gentry Citation1999; Boisserie et al. Citation2017a), A. crusafonti (Martino et al. Citation2021 and references therein), A. andrewsi (Pickford et al. Citation2022 and references therein), Hex. garyam (Boisserie et al. Citation2005), Hex. sivalensis (Falconer and Cautley Citation1836; de Visser Citation2008; Iqbal et al. Citation2019), Hex.? siculus (Martino et al. Citation2021 and references therein), Hex.? sahabiensis (Gaziry Citation1987; Pavlakis Citation2008), Hex.? hipponensis (Gaudry Citation1876) and S. mingoz (Boisserie et al. Citation2004).
Morphometric comparison
The dimensions of the remains from the Casino Basin are not easily comparable with other hippopotamid specimens. The P2 partially misses proximal and distal parts, and the maximum length is therefore underestimated. Isolated incisors are equally uninformative because their position cannot be deduced with certainty and thus compared with that of other hippopotamids. The dimensions of the incisors in the mandible are reported in and compared to different Late Miocene and earliest Pliocene hippopotamid species. However, the possible deformation of the true morphology does not permit a good morphometrical comparison. The second lower molar is almost complete and probably the best specimen collected from the Casino Basin. Nevertheless, the dimensions of the m2 are usually little diagnostic at specific level. (). It can only be concluded therefore that the specimen collected from the Casino Basin was a medium-size species of hippopotamine.
Table 2. Comparative dimensions of m2 (L: max length; W: max width) of Miocene hippopotamids, minimum (m)–maximum (M) dimensions in mm; mean in mm; number of specimens (N). Data for Hex.? siculus from Seguenza (Citation1902, Citation1907) and Hooijer (Citation1946); Hex.? sahabiensis from Gaziry (Citation1987) and Pavlakis (Citation2008); A. harvardi (1) from Coryndon (Citation1977) and Boisserie et al. (Citation2017a); A. harvardi (2) from Harrison (Citation1997); A. lothagamensis from Weston (Citation2000, Citation2003); A. qeshta from Boisserie et al. (Citation2017a); A. crusafonti from Faure and Méon (Citation1984) and Gentry and Hooker (Citation1988); A. andrewsi from Pickford et al. (Citation2022); Hex. garyam from Boisserie et al. (Citation2005); Hex. sivalensis from Hooijer (Citation1950) and de Visser (Citation2008); Hex. bruneti from Boisserie and White (Citation2004).
Table 3. Comparative dimensions of lower incisors (MD; LL) of Miocene hippopotamids (m–M dimensions in mm; mean in mm; N). Data for Hex.? siculus from Seguenza (Citation1902, Citation1907) and Hooijer (Citation1946); A. crusafonti from Faure and Méon (Citation1984) and Gentry and Hooker (Citation1988); Hex.? sahabiensis from Pavlakis (Citation2008); A. harvardi from Boisserie et al. (Citation2017a); A. lothagamensis from Boisserie et al. (Citation2017a); A. qeshta from Boisserie et al. (Citation2017a); Hex. garyam from Boisserie et al. (Citation2017a); Hex. sivalensis from Hooijer (Citation1950) and de Visser (Citation2008); Hex. bruneti from Boisserie and White (Citation2004).
Discussion
Characters of the Casino Basin hippopotamid in the framework of late Miocene hippopotamids variability
The hippopotamid remains collected from the Casino Basin are scarce and fragmentary. Nevertheless, some features can be highlighted. The Tuscan hippo displays an hexaprotodont condition probably with an i1 > i2 ≥ i3 arrangement, a c with finely crenulated and slightly transversally striated enamel with a groove on the mesial side, an m2 without accessory cuspids, well-developed cingulid on labial, mesial and distal sides, with a feebly trefoil wear pattern on the posterior cusps.
Among all the Late Miocene species, A. crusafonti is the earliest hippopotamid that shares a supposedly tetraprotodont condition, which is less archaic than the hexaprotodont one. This evolutionary trend is well observable in the different specimens of Hex. protamphibius collected from several strata with distinct ages in Shungura (Ethiopia): hippopotamids shifted from the hexaprotodont condition towards the more advanced tetraprotodont one (Coryndon Citation1978). However, the tetraprodont condition of A. crusafonti should probably be confirmed by further findings, as anomalies in the number of incisors have been extensively described in the literature (Falconer and Cautley Citation1868; Gaudry Citation1876; Stuenes Citation1989). The mandible collected from La Portera is currently under detailed studies since it was only described by Lacomba et al. (Citation1986) and never revised more recently. The i1> i2> i3 condition displays by the Casino Basin specimen is shared by A. harvardi (Weston Citation2003). In Hex. garyam the i2 is always the smallest one (Boisserie et al. Citation2005b). The overall aspect and dimensions of the incisors from the Casino Basin are different from the ones collected from the peri-Mediterranean area, Spain (A. crusafonti), Italy (Hex.? siculus), Egypt (A. andrewsi), Libya (Hex.? sahabiensis) and Algeria (Hex.? hipponensis) (Martino et al. Citation2021 and references therein; Pickford et al. Citation2022).
The c collected from the Casino Basin shows a distinctive structure of the enamel. The Casino basin specimen displays feeble transversal striae, finely crenulated enamel, and a longitudinal groove on the mesial side, while the c cross-section is D-shaped. Archaeopotamus crusafonti shares a c morphology really close to the one of the Casino Basin (pers. observ. R.M.). The finely crenulated enamel is also shared by A. harvardi, while Asian Hexaprotodon shows a great variability, from smooth to grooved enamel (Weston Citation2003; de Visser Citation2008). Hexaprotodon garyam shows an enamel smooth or finely striated (Boisserie et al. Citation2005). Hex.? siculus displayed two grooves on the medial side and one on the lateral side (Seguenza Citation1907). The cross-section in the c of the Casino Basin, even if partially incomplete, is mainly D-shaped with a groove on the mesial side. The c cross-section of A. crusafonti (MNCN62837) is similar to the one of the Casino Basin, mainly D-shaped with a groove on the mesial side, but probably more slightly mesial-labially compressed. A compressed canine in cross-section is displayed by A. lothagamensis, while other hippopotamids display more peculiar cross-section, pear-shaped, bean-shaped or D-shaped (Weston Citation2000, Citation2003; de Visser Citation2008; Boisserie et al. Citation2017a). The c cross-section of Hex. garyam is figured in Boisserie et al. (Citation2005b, p. 667-668, fig. 2A-3A) and it looks more rounded than in the Casino Basin specimen, A. lothagamensis and A. crusafonti. The cross-section of the c of most advanced Hippopotamidae is usually to triangular to D-shaped, while in more archaic forms the cross-section is more compressed (Weston Citation2000; Boisserie and White Citation2004). Kenyapotamus coryndonae, a basal Hippopotamidae, shows an ovate-triangular c cross-section (Tsujikawa Citation2005; Tsubamoto et al. Citation2015). Anthracotheriines in general have a c cross-section more rounded than hippopotamids (Boisserie et al. Citation2005a, Citation2010).
The m2 collected from the Casino Basin does not display prominent additional cuspids, and the cingulid is well visible. A. harvardi usually possesses four low cuspids with a prominent cingulid that forms shelves mesially and distally. Almost all Hexaprotodon m2s described in de Visser (Citation2008) show additional cuspids, heptaconid (present in 18 specimens out of 18 specimens analysed), pentaconid (15 out of 16) and hexaconid (13 out of 14). Cingulid is absent in most cases (16 out of 26), while rarely present on the lingual side (2 out of 26) and slightly more frequently on the labial side (7 out of 26). A similar lower molars complexity is also shared by Hex. palaeindicus (Falconer and Cautley Citation1868, p. 57, fig. 4a). In the light of the above-mentioned statements, the m2 from the Casino Basin is morphologically closer to Archaeopotamus rather than Asian Hexaprotodon representatives.
Paleobiogeographic considerations
Besides the Casino Basin, Tuscany is characterised by further Late Miocene-earliest Pliocene localities, such as Borro Strolla, Baccinello V3 and Velona (Rook et al. Citation1999; Ghetti et al. Citation2002; Rook and Martínez-Navarro Citation2004; Abbazzi et al. Citation2008; Pandolfi et al. Citation2021a, Citation2021b) (See Supplementary material, ). However, no hippopotamids have ever been reported from these Tuscan localities, which nonetheless present a similar faunal assemblage to the one described from the Casino Basin. The faunal list from Velona Basin (base of MN 13, ca. 7–6 Ma) includes Hipparion sp., Propotamochoerus sp., several cervids, bovids and Dipoides problematicus (Ghetti et al. Citation2002), while Baccinello V3 (6.733–6.436 Ma) comprises Pliorhinus megarhinus, Tapirus cf. arvernensis, Hippotherium malpassii, P. provincialis, bovids, several cervids and carnivorans (Rook et al. Citation1999; Rook and Martínez-Navarro Citation2004; Pandolfi and Rook Citation2017; Pandolfi et al. Citation2021a, Citation2021b, Citation2021c). The youngest locality, Borro Strolla (5.55–5.33 Ma) has a mammal list characterised by the occurrence of bovids, cervids and giraffids (Abbazzi et al. Citation2008; Pandolfi et al. Citation2021a, Citation2021b). In addition to Borro Strolla, Baccinello V3 and Velona, there are further Italian sites ascribed to Late Miocene: Verduno (late MN 13, 5.55–5.33 Ma), Moncucco Torinese (MN 13), Monticino Quarry (late MN 13, 5.55–5.33 Ma), Cessaniti (7.2–8.1 Ma) and Gravitelli (late MN13) (Seguenza Citation1902, Citation1907; Alessio et al. Citation1982; Gallai and Rook Citation2006, Citation2011; Angelone et al. Citation2011; Colombero et al. Citation2011; Marra et al. Citation2011, Citation2014; Rook et al. Citation2015, Citation2017, Citation2017; Pandolfi and Rook Citation2017; Pandolfi et al. Citation2019; Rook Citation2021; Pandolfi et al. Citation2021a, Citation2021b) (See Supplementary material, ). Verduno (Piedmont, west-northern Italy) is characterised by a rich mammal list, which encompasses a proboscidean, giraffids, a rhinocerotid, bovids, cervids, an equid and several carnivorans (Colombero et al. Citation2014). Moncucco Torinese (Piedmont, west-northern Italy) faunal list comprises Mesopithecus sp., a rhinocerotid, Tapirus arvernensis, cervids, bovids and several carnivorans (Angelone et al. Citation2011; Colombero et al. Citation2017), while fossil mammals from Monticino Quarry (Emilia Romagna, central-northern Italy) include M. pentelicus, a proboscidean, H. malpassii, a rhinocerotid, P. provincialis, an aardvark, a honey badger, bovids, and several carnivorans (Rook et al. Citation2015; Rook Citation2021; Pandolfi et al. Citation2021a, Citation2021b). Cessaniti (Calabria, South Italy) has a faunal list, which comprises giraffids, a rhinocerotid, a proboscidean (Marra et al. Citation2011, Citation2017). Hippopotamid presence from this locality can be for now ruled out because most likely the specimens from Cessaniti (an incisor and a femur) belong to an anthracotheriid (Marra et al. Citation2017). Gravitelli (Sicily, South Italy) documented the presence of Mesopithecus sp., carnivorans, proboscideans, a rhinocerotid, P. provincialis. and several artiodactyls, including Hex.? siculus (Seguenza Citation1902; Citation1907; Gallai and Rook Citation2006; Pandolfi and Rook Citation2017; Martino et al. Citation2021; Pandolfi et al. Citation2019, Citation2021a, Citation2021b; Iannucci Citation2023). All the above-mentioned Italian localities display mammals’ assemblages richer than the Casino Basin one including carnivorans, giraffids, different bovids and cervids species, proboscideans, equids and rhinocerotids. These large mammals were unearthed from different peri-Mediterranean localities in association with hippopotamids (Gravitelli, Venta del Moro, El Arquillo I, Las Casiones, As Sahabi, Wadi Natrun) (Seguenza Citation1902, Citation1907; Alcalá Citation1994; Alcalá and Montoya Citation1998; Bernor and Rook Citation2008; Morales et al. Citation2011; Pickford et al. Citation2022) (see Supplementary material). In particular, the absence of proboscideans and rhinoceros, as well as giraffids, probably suggests a ‘barrier’ to the dispersion of these mammals in the Casino Basin.
Among all the Late Miocene localities of the peri-Mediterranean area where hippopotamids occurred (Spanish sites, Gravitelli, As Sahabi, Point-De-Duvivier and Wadi Natrun all reported in ), the mammal assemblage of the Casino basin more closely resembles the fauna unearthed from Spain, in particular from Venta del Moro (Morales et al. Citation2011 and references therein) and El Arquillo I (Alcalá and Montoya Citation1998), which include D. problematicus, P. provincialis, Thalassictis, Hipparion, Parabos and Eucyon.
The peculiar mosaic characteristics of Hex.? sahabiensis should be better investigated in order to disclose if this species might be the ancestor of the Late Miocene European ones. As already pointed out by Weston (Citation2000), some characters are close to Archaeopotamus. However, the sagittal section of the mandibular symphysis does not resemble the one of Archaeopotamus (Pavlakis Citation2008; Boisserie et al. Citation2017a). Hexaprotodon? sahabiensis should be therefore revised in order to disclose a possible massive event of dispersion of Archaeopotamus from the Central Africa-Arabian Peninsula to North Africa (As Sahabi and Wadi Natrun) and Europe (Casino Basin, Iberian Peninsula and France) (Gaziry Citation1987; Pavlakis Citation2008; Martino et al. Citation2021 and references therein; Pickford et al. Citation2022). The closeness of the Casino Basin fauna with the Spanish ones, where A. crusafonti is well documented, can support a dispersal event in the Tuscan area through the Iberian Peninsula. In that regard, Ghetti et al. (Citation2002) argued for a full land connection between Tuscany and the European mainland during the Late Miocene, which may therefore have provided an easy dispersal way for mammals. The same connection between Central Italy and the European mainland around 6.3 Ma (M1-2 transition) is also highlighted by Van der Made et al. (Citation2006) (). Rögl (Citation1999), Ferretti et al. (Citation2003), Cirrincione et al. (Citation2015) and Broquet (Citation2016) postulated that the areas of Sicily and Calabria were close to North Africa during the Tortonian. Van der Made et al. (Citation2006) hypothesised two different scenarios regarding South Italy: (i) Calabria was part of the African land mass or (ii) it was temporarily connected during a regression. A dispersal from Calabria to North during the Messinian may have been therefore prevented due different paleoenvironmental conditions inferred from the fauna or because Calabria was separated from northern areas. The other possible dispersal route, from North Africa through Sicily up to the Casino Basin, should be better investigated. Nevertheless, the partial isolation of the Tusco-Sardinian paleobioprovince with the Calabro-Sicilian paleobioprovince during the Late Miocene would not have probably allowed the dispersal of hippopotamids from Sicily, where Hex.? siculus is reported (Rook et al. Citation2006). Hippopotamids are currently unknown from Late Miocene Eastern European sites, and their presence in Greece is doubtful, therefore a Balkan dispersal event can be discarded as well (Athanassiou Citation2022).
Conclusions
Most of the peri-Mediterranean hippopotamids are poorly known, and most of them are dubiously ascribed to the genus Hexaprotodon. Hexaprotodon? sahabiensis, Hex.? siculus, Hex.? hipponensis, Archaeopotamus andrewsi and A. crusafonti display characters that are not shared by the Casino Basin specimens, which is characterised by finely crenulated c enamel, with subtly transversal lines and a groove on the mesial side that is less defined than in other Late Miocene species. The c cross-section is D-shaped. The incisors are conical and straight and in cross-sections are mesial-laterally compressed. The m2 is simply built, cingulid is well developed on mesial, distal and labial sides, accessory cuspids are absent and the trefoil wear pattern is developed on the posterior cusps. In the light of these characters, the species erected by Joleaud (Citation1920), originally named as Hippopotamus pantanellii, may be considered valid. The species collected from the Casino Basin is here reassigned, with caution due to their fragmentary nature, to Archaeopotamus.
The characters shared by Tuscan specimens in particular, the m2 without accessory cuspids and with cingulid on labial, mesial and distal sides, together with the i1> i2> i3 configuration, permit to exclude the genus Hexaprotodon. In Hex. garyam the i2 is always the smallest incisor, while in A. pantanellii is the i3. In the Asian lineage of Hexaprotodon, the m2 is fairly more complex than the one collected from the Casino basin. The c and m2 of A. crusafonti, poorly described in literature, should be better investigated to evaluate a possible synonymy with the Tuscan species. In case of synonymy with A. crusafonti, the Casino Basin species A. pantanellii would have the priority.
The occurrence of Archaeopotamus in the Casino Basin is very significant. The clear African affinity of this taxon testifies that the Late Miocene Hippopotamidae probably dispersed from the North Africa to Europe via the Iberian Peninsula. A dispersal event from Sicily does not seem likely, given the suggested palaeogeography of Italy during the Late Miocene, as well as a dispersion via the Balkans, because, hippopotamids have never been reported from that area.
Supplemental Material
Download MS Word (23.7 KB)Supplemental Material
Download MS Excel (14.8 KB)Acknowledgments
This research benefited from the grant UIDB/04035/2020 by the Fundação para a Ciência e Tecnologia to GeoBioTec. R.M. wants to thank Prof. Martin Pickford for the help and for the fruitful conversations. The authors also thank C. Dal Sasso and G.G. Bardelli (MSNM), E. Cioppi (IGF), R. Barbieri (MGGC), C. Mangoni (MCZR), C. Sarti (MSNAF), F. Farsi, G. Manganelli (MZUF) and C. Sorbini (MSNCC) for their assistance and support during the visits to fossil collections. This research received support from the SYNTHESYS+ project http://www.synthesys.info/, which is financed by the European Community Research Infrastructure Action under the H2020 Integrating Activities Programme, Project number 823827 at the Museo Nacional de Ciencias Naturales (CSIC). L.P. thanks the European Commission’s Research Infrastructure Action, EU-SYNTHESYS+ project BE-TAF-1367; part of this research received support from the SYNTHESYS+ Project http://www.synthesys.info/which is funded by the European Commission.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/08912963.2023.2194912
Additional information
Funding
References
- Abbazzi L. 2001. Cervidae and Moschidae (Mammalia, Artiodactyla) from the Baccinello V-3 assemblage (Late Miocene, Late Turolian, Grosseto, Italy). Riv Ital Paleontol. 107(1):106–123.
- Abbazzi L, Delfino M, Gallai G, Trebini L, Rook L. 2008. NEW DATA ON THE VERTEBRATE ASSEMBLAGE OF FIUME SANTO (NORTH-WEST SARDINIA, ITALY), AND OVERVIEW ON THE LATE MIOCENE TUSCO-SARDINIAN PALAEOBIOPROVINCE. Palaeontology. 51(2):425–451. doi:10.1111/j.1475-4983.2008.00758.x.
- Aguirre ED. 1963. Hippopotamus crusafonti n. sp. del Plioceno inferior de Arenas del Rey (Granada). Notas y comunicaciones del Instituto Geológico y Minero de España. 69:215–230.
- Aguirre ED, Robles F, Thaler L, López Martínez N, Alberdi MT, Fuentes C. 1973. Venta del Moro, nueva fauna finimiocena de Moluscos y Vertebrados. Estud Geol. 29:569–578.
- Alcalá L. 1994. Macromamíferos neógenos de la fosa de Alfambra-Teruel. Madrid: Instituto de Estudios Turolenses-Museo Nacional de Ciencias Naturales-CSIC; p. 554.
- Alcalá L, Montoya P. 1998. Hexaprotodon crusafonti (Hippopotamidae, Mammalia) del Mioceno superior de El Arquillo (Fosa de Teruel). Bol R Soc Esp Hist Nat, Secc Geol. 94:93–100.
- Alessio M, Allegri L, Ambrosetti P, Bartolomei G, Bella F, Belluomini G, Calderoni G, Carraro F, Charrier G, Cortesi C, et al. 1982. Il giacimento fossilifero pleistocenico superiore di Moncucco Torinese. Geogr Fis Dinam Quat. 5:219–239.
- Andrews CW. 1902. Note on a Pliocene vertebrate fauna from the Wadi-Natrun, Egypt. Geol Mag. 9:433–439. doi:10.1017/S0016756800178112.
- Angelone C, Colombero S, Daniela E, Piero G, Federica M, Pavia M, Trenkwalder S, Van Den Hoek Ostend LW, Zunino M, Pavia G. 2011. Moncucco Torinese, a new post-evaporitic Messinian fossiliferous site from Piedmont (NW Italy). Neues Jahrb Geol Palaontol Abh. 259(1):89–104. doi:10.1127/0077-7749/2010/0108.
- Arambourg C. 1944. Au sujet de l’Hippopotamus hipponensis Gaudry. Bulletin de la Société Géologique de France S5-XIV(4–6):147–152. doi:10.2113/gssgfbull.S5-XIV.4-6.147.
- Arambourg C. 1947. Contribution a l’étude géologique et paléontologique du bassin du lac Rudolphe et de la basse vallée de l’Omo. Mission scientifique de l’Omo 1932–1933. Géologie-Anthropologie. 1:232–562.
- Athanassiou A. 2022. The Fossil Record of Continental Hippopotamids (Mammalia: Artiodactyla: Hippopotamidae) in Greece. In: Vlachos E, editor. Fossil Vertebrates of Greece (Vol. 2). Cham: Springer; p. 281–299.
- BERNOR R.L. & ROOK L. 2008. A current view of Sahabi large mammal biogeographic relationships. In Boaz N.T., El-Arnauti A, Pavlakis P. and Salem M., (Editor(s). Circum-Mediterranean Neogene Geology and Biotic Evolution: The Perspective from Libya. Garyounis Scientific Bulletin. Special Issue, no 5; 285–292.
- Boisserie JR. 2007. Family Hippopotamidae. In: Prothero DR, Foss SE, editors. The Evolution of Artiodactyls. Baltimore: Johns Hopkins University Press; p. 106–119.
- Boisserie JR, Bibi F. 2022. Hippopotamidae from the Baynunah Formation. In: Bibi F, Kraatz B, Beech M, Hill A, editors. Sands of Time: ancient life in the late Miocene of Abu Dhabi, United Arab Emirates. Cham: Springer International Publishing; p. 243–260.
- Boisserie J-R, Brunet M, Andossa L, Vignaud P. 2003. Hippopotamids from the Djurab Pliocene faunas, Chad, Central Africa. J African Earth Sci. 36(1–2):15–27. doi:10.1016/S0899-5362(03)00021-6.
- Boisserie J-R, Lihoreau F, Brunet M. 2005a. Origins of Hippopotamidae (Mammalia, Cetartiodactyla): towards resolution. Zool Scr. 34(2):119–143. doi:10.1111/j.1463-6409.2005.00183.x.
- Boisserie J-R, Lihoreau F, Orliac M, Fisher RE, Weston EM, Ducrocq S. 2010. Morphology and phylogenetic relationships of the earliest known hippopotamids (Cetartiodactyla, Hippopotamidae, Kenyapotaminae). Zool J Linn Soc. 158(2):325–366. doi:10.1111/j.1096-3642.2009.00548.x.
- Boisserie JR, Likius A, Vignaud P, Brunet M. 2005b. A new late Miocene hippopotamid from Toros-Ménalla, Chad. J Vertebr Paleontol. 25(3):665–673. doi:10.1671/0272-4634(2005)025[0665:ANLMHF]2.0.CO;2.
- Boisserie J-R, Schuster M, Beech MJ, Hill A, Bibi F. 2017a. A new species of hippopotamine (Cetartiodactyla, Hippopotamidae) from the late Miocene Baynunah Formation, Abu Dhabi, United Arab Emirates. Palaeovertebrata. 41(1):e2. doi:10.18563/pv.41.1.e2.
- Boisserie J-R, Suwa G, Asfaw B, Lihoreau F, Bernor RL, Katoh S, Beyene Y. 2017b. Basal hippopotamines from the upper Miocene of Chorora, Ethiopia. J Vertebr Paleontol. 37(3):e1297718. doi:10.1080/02724634.2017.1297718.
- Boisserie J-R, White TD. 2004. A new species of Pliocene Hippopotamidae from the Middle Awash, Ethiopia. J Vertebr Paleontol. 24(2):464–473. doi:10.1671/2475.
- Bossio A, Mazzei R, Salvatorini G, Sandrelli F. 2002. Geologia dell’area compresa tra Siena e Poggibonsi (“Bacino del Casino”). Atti Soc Toscana Sci Nat Mem A. 107:69–85.
- Broquet P. 2016. Sicily in its Mediterranean geological frame. Boletin Geoologico Y Min. 127:547–562.
- Capellini G. 1872. Atti della Sesta Riunione straordinaria della Società italiana di Scienze Naturali tenuta in Siena nei giorni 22, 23, 24 Settembre 1872. Atti Società Italiana Scienze Naturali. 15:190–216.
- Cirilli O, Bernor RL, Rook L. 2020. New Evidences on the Hipparion sample from Il Casino (Siena, Late Miocene, Italy). Preliminary review. Fossilia. 2020:9–11. doi:10.32774/FosRepPal.2020.0603.
- Cirrincione R, Fazio E, Fiannacca P, Ortolano G, Pezzino A, Punturo R. 2015. The Calabria-Peloritani Orogen, a composite terrane in Central Mediterranean; its overall architecture and geodynamic significance for a pre-Alpine scenario around the Tethyan Basin. Period Di Mineral. 84:701–749.
- Colombero S, Alba DM, D’Amico C, Delfino M, Esu D, Giuntelli P, Harzhauser M, Mazza PPA, Mosca M, Neubauer TA, et al. 2017. Late Messinian mollusks and vertebrates from Moncucco Torinese, north-western Italy. Paleoecological and Paleoclimatological Implications. Palaeontol Electron. 20(1.10A):1–66.
- Colombero S, Angelone C, Bonelli E, Carnevale G, Cavallo O, Delfino M, Giuntelli P, Mazza P, Pavia G, Pavia M. 2014. The upper Messinian assemblages of fossil vertebrate remains of Verduno (NW Italy): another brick for a latest Miocene bridge across the Mediterranean. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 272(3):287–324. doi:10.1127/0077-7749/2014/0408.
- Colombero S, Angelone C, Pavia M. 2011. Preliminary data on fossil rodents from the vertebrate sites Moncucco and Verduno (post-evaporitic Messinian, Piedmont, NW Italy). In: Carnevale G, Patacca E, Scandone P, editors. CMNS Interim Colloquium, Neogene Park 1-3. Scontrone (L’Aquila): Abstract Book; p. 25–27.
- Coryndon SC. 1977. The taxonomy and nomenclature of the Hippopotamidae (Mammalia, Artiodactyla) and a description of two new fossil species. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen; Amsterdam. 80:61–88.
- Coryndon SC. 1978. Hippopotamidae. In: Maglio VJ, Cooke HBS, editors. Evolution of African Mammals. Cambridge: Harvard University Press; p. 483–495.
- Crusafont M, Adrover R, Golpe JM. 1964. Découverte dans le Pikermien d’Espagne du plus primitif des hippopotames: hippopotamus (Hexaprotodon) primaevus n. sp. C R Hebd Séances Acad Sci. 258:1572–1575.
- de Visser J. 2008. The Extinct Genus Hexaprotodon Falconer & Cautley, 1836 (Mammalia, Artiodactyla, Hippopotamidae) in Asia: paleoecology and Taxonomy. Unpublished, PhD thesis. Universiteit Utrecht; p 390.
- Falconer H, Cautley PT. 1836. Note on the fossil Hippopotamus of the Siwalik hills. Asiatic Researches. 19:39–53.
- Falconer H, Cautley PT. 1868. Introductory observations on the geography, geological structure and fossil remains in the Siwalik hills. Palaeontological Memoirs. 1:1–29.
- Faure M, Méon H. 1984. L’Hippopotamus crusafonti de La Mosson (près Montpellier). Première reconnaissance d’un Hippopotame néogène en France. Comptes-rendus des séances de l’Académie des sciences. Série 2, Mécanique-physique, chimie, sciences de l’univers. Sciences de la Terre. 298(3):93–98.
- Ferretti MP, Rook L, Torre D. 2003. Stegotetrabelodon (Proboscidea, Elephantidae) from the late Miocene of southern Italy. J Vertebr Paleontol. 23(3):659–666. doi:10.1671/2353.
- Forsyth Mayor CI. 1875. Considerazioni sulla fauna dei Mammiferi pliocenici e postpliocenici della Toscana. Memorie Società Toscana Scienze Naturali. 1:223–245.
- Gallai G, Rook L. 2006. Propotamochoerus sp. (Suidae, Mammalia) from the late Miocene Gravitelli (Messina, Sicily, Italy) rediscovered. Riv Ital Paleontol. 112(2):317–321.
- Gallai G. 2005. Tafonomia e paleobiologia della fauna mammaliana turoliana (Miocene superiore) del bacino del Casino (Siena): risultati preliminari. Rendiconti Società Paleontologica Italiana. Riv Ital Paleontol. 2:119–125.
- Gallai G, Rook L. 2011. Propotamochoerus provincialis (Gervais, 1859) (Suidae, Mammalia) from the latest Miocene (late Messinian; MN13) of Monticino Quarry (Brisighella, Emilia-Romagna, Italy). Boll Soc Paleontol Ital. 50(1):29–34.
- Gaudry A. 1876. Sur un Hippopotame fossile découvert à Bone (Algérie). BSGF-EARTH SCI B. 4(3):502–504.
- Gaziry AW. 1987. Hexaprotodon sahabiensis (Artiodactyla, Mammalia): a new hippopotamus from Libya. In: Boaz NT, El-Arnauti A, Gaziry AW, de Heinzelin J, Boaz D, editors. Neogene Paleontology and Geology of Sahabi. New York: Alan R. Liss; p. 303–315.
- Gentry AW. 1999. A fossil hippopotamus from the Emirate of Abu Dhabi, United Arab Emirates. In: Whybrow PJ, Hill A, editors. Fossil vertebrates of Emirati Arabi. New Haven: Yale University Press; p. 271–289.
- Gentry A, Hooker J. 1988. The phylogeny of the Artiodactyla. In: Benton M, editor. The Phylogeny and Classification of the Tetrapods (Vol. 2). Oxford: Clarendon Press; p. 235–272.
- Geraads D, Reed D, Barr WA, Bobe R, Stamos P, Alemseged Z. 2021. Plio-Pleistocene mammals from Mille-Logya, Ethiopia, and the post-Hadar faunal change. Journal of Quaternary Science. 36(6):1073–1089. doi:10.1002/jqs.3345.
- Gèze R. 1985. Répartition paléoécologique et relations phylogénétiques des Hippopotamidae (Mammalia, Artiodactyla) du néogéne d'Afrique Orientale. In: Beden M, Behrensmeyer AK, Boaz NT, Bonnefille R, Brain CK, Cooke B, Coppens Y, Dechamps R, Eisenmann V, Gentry A, Geraads D, Gèze R, Guérin C, Harris J, Koeniguer J, Letouzey F, Petter G, Vincens A, Vrba E, editor(s). L'environnement des hominidés au Plio-Pléistocène. Paris: Fondation Singer-Polignac-Masson; p. 81–100.
- Ghetti P, Anadón P, Bertini A, Esu D, Gliozzi E, Rook L, Soulié-Märsche I. 2002. The Early Messinian Velona Basin (Siena, Central Italy): paleoenvironmental and paleobiogeographical reconstructions. Palaeogeogr Palaeoclimatol Palaeoecol. 187(1–2):1–33. doi:10.1016/S0031-0182(02)00503-5.
- Gray JE. 1821. On the natural arrangement of vertebrate animals. London Medical Repository. 15:296–310.
- Harris JM, Leakey MG, Cerling TE, Winkler AJ. 2003. Early Pliocene tetrapod remains from Kanapoi, Lake Turkana Basin, Kenya. Contrib Sci. 498:39–114. doi:10.5962/p.226790.
- Harrison T. 1997. The anatomy, paleobiology, and phylogenetic relationships of the Hippopotamidae (Mammalia, Artiodactyla) from the Manonga Valley, Tanzania. In: Harrison T, editor. Neogene Paleontology of the Manonga Valley, Tanzania. New York: Plenum Press; p. 137–190.
- Hooijer DA. 1946. Notes On Some Pontian Mammals From Sicily, Figured By Seguenza. Archives néerlandaises de Zoologie. 7(3):301–333. doi:10.1163/036551646X00031.
- Hooijer DA. 1950. The fossil Hippopotamidae of Asia, with notes on the Recent species. Zool. Verh. 8:1–124.
- Iannucci A. 2023. A reappraisal of the lost suids from the Late Miocene of Gravitelli (Sicily, Italy) and paleobiogeographical implications. Palaeoworld in press Palaeoworld. 10.1016/j.palwor.2023.02.001
- Iannucci A, Cherin M, Sorbelli L, Sardella R. 2021. Suidae transition at the Miocene-Pliocene boundary: a reassessment of the taxonomy and chronology of Propotamochoerus provincialis. Journal of Mammalian Evolution. 28(2):323–335. doi:10.1007/s10914-020-09521-w.
- Iqbal A, Khan AM, Ahmad RM, Khan MA, Akhtar M. 2019. Systematics and Palaeo-Environmental Implications of Hexaprotodon sivalensis (Hippopotamidae, Mammalia) from the Plio-Pleistocene Upper Siwaliks (Pakistan). In: Boughdiri M, editor. Paleobiodiversity and Tectono-Sedimentary Records in the Mediterranean Tethys and Related Eastern Areas, Advances in Science, Technology & Innovation. Switzerland: Springer Nature; p. 65–67.
- Joleaud L. 1920. Contribution à l’étude des hippopotames fossiles. BSGF-EARTH SCI B. 4(20):13–26.
- Lacomba JI, Morales J, Robles F, Santiesteban C, Alberdi MT. 1986. Sedimentología y paleontología del yacimiento Finimioceno de La Portera (Valencia). Estudios Geológicos. 42(2–3):167–180. doi:10.3989/egeol.86422-3746.
- Lazzarotto A, Sandrelli F. 1977. Stratigrafia e Assetto Tettonico delle Formazioni Neogeniche nel Bacino del Casino (Siena). Boll Soc Geol Ital. 96:747–762.
- Linnaeus C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Stockholm: Laurentii Salvii; p. 824. Editio decima, reformata
- Major CF. 1874. Considerazioni sulla fauna de mammiferi plioceni e postpliocenici della Toscana. Atti Soc Tosc Sci Nat. 1(1):8–40.
- Major CF. 1877. Considerazioni sulla Fauna dei Mammiferi pliocenici e post-pliocenici della Toscana. Atti Soc Tosc Sci Nat. 1:207–227.
- Marra AC, Carone G, Agnini C, Ghinassi M, Oms O, Rook L. 2017. Stratigraphic and chronologic framework of the upper Miocene Cessaniti succession (Vibo Valentia, Calabria, Italy). Riv Ital Paleontol. 123(3):379–393.
- Marra AC, Solounias N, Carone G, Rook L. 2011. Palaeogeographic significance of the giraffid remains (Mammalia, Arctiodactyla) from Cessaniti (Late Miocene, Southern Italy). Géobios. 44:189–197.
- Martino R, Pignatti J, Rook L, Pandolfi L. 2020. Systematic revision of hippopotamid remains from the Casino Basin, Tuscany, Italy. Fossilia. 2020:29–31.
- Martino R, Pignatti J, Rook L, Pandolfi L. 2021. Hippopotamid dispersal across the Mediterranean in the latest Miocene: a re-evaluation of the Gravitelli record from Sicily, Italy. Acta Palaeontol Pol. 66(3):s067–s078.
- Mazza PPA. 1995. New evidence on the Pleistocene hippopotamuses of western Europe. Geol Rom. 31:61–241.
- Mbua E, Kusaka S, Kunimatsu Y, Geraads D, Sawada Y, Brown FH, Sakai T, Boisserie JR, Saneyoshi M, Omuombo C, et al. 2016. Kantis: a new Australopithecus site on the shoulders of the Rift Valley near Nairobi, Kenya. J Hum Evol. 94:28–44.
- McDougall IAN, Brown FH. 2008. Geochronology of the pre-KBS tuff sequence, Omo Group, Turkana Basin. J Geol Soc London. 165(2):549–562.
- McDougall IAN, Feibel CS. 1999. Numerical age control for the Miocene-Pliocene succession at Lothagam, a hominoid-bearing sequence in the northern Kenya Rift. J Geol Soc London. 156(4):731–745.
- Montgelard C, Catzeflis FM, Douzery E. 1997. Phylogenetic relationships of artiodactyls and cetaceans as deduced from the comparison of cytochrome b and 12S rRNA mitochondrial sequences. Mol Biol Evol. 14(5):550–559.
- Morales J. 1984. Venta del Moro: su Macrofauna de Mamiferos y Biostratigrafia continental del Mioceno terminal mediterraneo. Unpublished PhD thesis. Univ. Complutense de Madrid; p. 340.
- Morales J, Montoya P, Abella J. 2011. Venta del Moro: un yacimiento clave para conocer la historia del lìmite Mio-Plioceno en el area mediterranea. Isurus. 4:60–71.
- Morton SG. 1844. On a supposed new species of Hippopotamus. Proceedings of the National Academy of Sciences, Philadelphia. 2:4–27.
- Pandolfi L, Marra AC, Carone G, Maiorino L and Rook L. (2019). A new rhinocerotid (Mammalia, Rhinocerotidae) from the latest Miocene of Southern Italy. Hist Biol, 33(2), 194–208. 10.1080/08912963.2019.1602615.
- Pandolfi L, Masini F, Kostopoulos DS. 2021a. Messinian Italian Bovidae revised: paleobiogeographic and biochronological implications. Hist Biol. 33(12):3590–3603.
- Pandolfi L, Masini F, Kostopoulos DS. 2021b. The latest Miocene small-sized Cervidae from Monticino Quarry (Brisighella, Central Italy): paleobiogeographic and biochronological implications. Hist Biol. 33(12):3368–3374.
- Pandolfi L, Pierre-Olivier A, Bukhsianidze M, Lordkipanidze D and Rook L. (2021c). Northern Eurasian rhinocerotines (Mammalia, Perissodactyla) by the Pliocene–Pleistocene transition: phylogeny and historical biogeography. J Syst Palaeontol, 19(15), 1031–1057. 10.1080/14772019.2021.1995907.
- Pandolfi L, Rook L. 2017. Rhinocerotidae (Mammalia, Perissodactyla) from the latest Turolian localities (MN 13; late Miocene) of central and northern Italy. Boll Soc Paleontol Ital. 56(1):45–56.
- Pantanelli D. 1879. Sugli strati miocenici del Casino (Siena) e considerazioni sul Miocene superiore.Atti della Reale Accademia dei Lincei, Memorie della classe di scienze fisiche, matematiche e naturali. 3(3):309–327.
- Pantanelli D. 1886. Monografia degli strati pontici del Miocene superiore nell’Italia settentrionale e centrale. Memorie della R. Accademia di Scienze, Lettere ed Arti di Modena, Sezione di Scienze. 4:127–231.
- Pavlakis P. 2008. Rediscovered hippopotamid remains from As Sahabi. Circum-Mediterranean geology and biotic evolution during the Neogene period: the perspective from Libya. Garyounis Scientific Bulletin. (5):179–187.
- Pickford M, Abdel Ghany MS, Sileem AH, Gamil MM. 2022. A descriptive catalogue of fossils from Wadi Natrun (Mio-Pliocene) Egypt, housed in the Cairo Geological Museum and in Museums in Europe. München: Münchner Geowissenschaftliche Abhandlungen Reihe A. Geologie und Paläontologie, Verlag Dr. Friedrich Pfeil; p. 92.
- Popov SV, Rögl F, Rozanov AY, Steininger FF, Shcherba IG, Kovac M. 2004. Lithological-paleogeographic maps of Paratethys-10 maps late Eocene to Pliocene. Cour. Forsch. Inst. Senckenberg. 250:1–46.
- Rögl F. 1999. Mediterranean and paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview). Geol Carpath. 50:330–349.
- Rook L. 1992. Italian Messinian localities with vertebrate faunas. Paleontologia i evolució. 24(25):141–147.
- ROOK L, editor. 2021. La fauna messiniana di Cava Monticino. Memorie dell’Istituto Italiano di Speleologia. Brisighella, RA, (Serie II; Vol. 37, p. 1–210).
- Rook L, Abbazzi L, Engesser B. 1999. An overview on the Italian Miocene land mammal faunas. In: In: Agustı́ J, Rook L, Andrews P, editors. The Evolution of Neogene Terrestrial Ecosystems in Europe. Cambridge: University Press; p. 191–204.
- Rook L, Bernor RL. 2013. Hippotherium malpassii (Equidae, Mammalia) from the latest Miocene (late Messinian; MN13) of Monticino gypsum quarry (Brisighella, Emilia-Romagna, Italy). Boll Soc Paleontol Ital. 52:95–102.
- Rook L, Delfino M, Sami M. 2015 I vertebrati fossili della Cava del Monticino di Brisighella: una finestra sui popolamenti continentali del Mediterraneo nel Miocene Superiore. In: Piastra, S, Lucci, P, editor(s). I Gessi di Brisighella e Rontana-Castenuovo, Memorie dell’Istituto italiano di Speleologia. (S. II; vol. 28). 79–100.
- Rook L, Gallai G, Torre D. 2006. Lands and endemic mammals in the Late Miocene of Italy: constrains for paleogeographic outlines of Tyrrhenian area. Palaeogeogr Palaeoclimatol Palaeoecol. 238(1–4):263–269.
- Rook L, Martínez-Navarro B. 2004. Viverra howelli n. sp., a new viverrid (Carnivora, Mammalia) from the Baccinello–Cinigiano Basin (latest Miocene, Italy). Riv. Ital. Paleontol. 110(3):719–723.
- Rook L, Mazza P, Rustioni M, Torre D. 2000. Lands and endemic mammals in the Late Miocene of Italy: paleogeographic outlines of Tyrrhenian and Adriatic areas between 11–9 and 7– 4 Ma. Lyon, France: ESF/EEDEN ESF Scientific Programme on “Environments and Ecosystem Dynamics of the Eurasian Neogene” (EEDEN). Workshop on “State-of-the-art; Lyon”; p. 56–58.
- Rütimeyer L. 1876. Über Pliocen und Eisperiode auf beiden Seiten der Alpen. Basel: Georg; p. 76.
- Seguenza G. 1902. I vertebrati fossili della Provincia di Messina. Parte seconda. Mammiferi e geologia del Piano Pontico. Boll Soc Geol Ital. 21:115–175.
- Seguenza G. 1907. Nuovi resti di mammiferi pontici di Gravitelli presso Messina. Boll Soc Geol Ital. 26:89–122.
- Stehlin HG. 1899-1900. Über die Geschichte des Suiden Gebisses. Abhandlungen der Schweizerischen. Zürich: Paläontologischen Gesellschaft; p. 527.
- Stromer E. 1914. Mitteilungen über Wirbeltierreste aus dem Mittelpliocän des Natrontales (Ägypten), 3. Artiodactyla: a. Bunodontia: flußpferd. Zeitschrift der Deutschen Geologischen Gesellschaft. 66:1–33.
- Stuenes S. 1989. Taxonomy, habits, and relationships of the subfossil Madagascan hippopotami Hippopotamus lemerlei and H. madagascariensis. J Vertebr Paleontol. 9(3):241–268.
- Thenius E. 1989. Zähne und Gebiß der Säugetiere. Handbuch der Zoologie. De Gruyter: Berlin. Vol. VIII(56): 513.
- Tsubamoto T, Yutaka K, Nakaya H, Sakai T, Saneyoshi M, Mbua E, Nakatsukasa M. 2015. New specimens of a primitive hippopotamus, Kenyapotamus coryndonae, from the Upper Miocene Nakali Formation, Kenya. J Geol Soc Japan. 121(4):153–159.
- Tsujikawa H. 2005. The updated late Miocene large mammal fauna from Samburu Hills, northern Kenya. Afr Study Monogr. (32):1–50.
- Van der Made J, Morales J, Montoya P. 2006. Late Miocene turnover in the Spanish mammal record in relation to palaeoclimate and the Messinian Salinity Crisis. Palaeogeogr Palaeoclimatol Palaeoecol. 238(1–4):228–246.
- Weston EM. 2000. A new species of hippopotamus Hexaprotodon lothagamensis (Mammalia: Hippopotamidae) from the late Miocene of Kenya. J Vertebr Paleontol. 20:177–185.
- Weston EM. 2003. Fossil Hippopotamidae from Lothagam. In: Leakey MG, Harris JM, editors. Lothagam. The Dawn of Humanity in Eastern Africa. New York: Columbia University Press; p. 441–483.