Figures & data
Figure 1. Components of gut microbiota, their approximate size and abundance.
The gut microbiota includes viruses, archaea, bacteria, fungi, and parasites. These diverse microorganisms vary in size and abundance in the gut. Only approximate numbers are given for the colon; absolute abundance can vary by orders of magnitude based on location in the gut and applied analysis methods.Citation31,Citation225
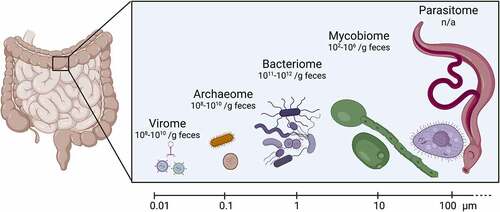
Figure 2. Effect of the gut virome on enteric bacterial infections.
Viruses in the gut are primarily bacteriophages and eukaryotic viruses. (1) Prophages alter bacterial phenotypes by encoding genes involved in antibiotic resistance, virulence factors, toxins and regulation of virulence factors. For example, the S. Typhimurium prophage Sopɸ encodes for SopE, which is required for invading the intestinal epithelium. Prophage Gifsy-1 encodes SodC and GipA, which are required for survival in macrophages and Peyer’s patches, respectively. (2) Bacteriophages can induce bacterial lysis, releasing pathogen-associated molecular patterns (PAMPs). These bacterial PAMPS are recognized by antigen-presenting cells (APCs) via pathogen recognition receptors, such as TLRs, leading to the release of proinflammatory cytokines. (3) Bacteriophages can also coat the mucus layer of the gut, preventing bacterial binding to the epithelium. (4) Eukaryotic RNA viruses are detected by cytosolic viral RNA-sensing receptor, RIG-I, resulting in the release of IL-15. IL-15 helps to maintain the number of intestinal intraepithelial lymphocytes (IELs), strengthening the intestinal defense against bacterial infection. (5) Nucleic acids of gut eukaryotic viruses are sensed by TLR3 and TLR7 on APCs, which induces secretion of anti-inflammatory interferon β (IFN-β). (6) Eukaryotic viruses such as murine norovirus induce an interleukin-23 (IL-23)-dependent burst of IL-22 secretion by innate lymphoid cells. The resulting enhanced production of the antimicrobial peptide Reg3γ by epithelial cells can protect against infection by Citrobacter rodentium and Vancomycin-resistantEnterococcus.
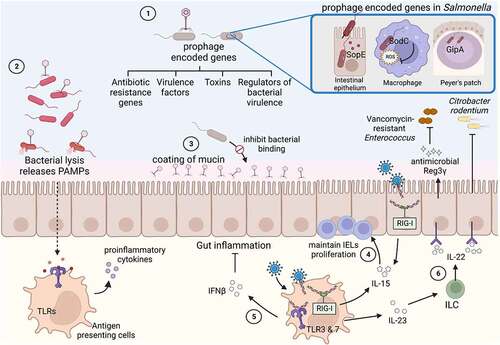
Figure 3. Effect of the gut archaeome on enteric bacterial infections.
Methanogenic archaea dominate the archaeome in the human gut. Bacteria ferment dietary carbohydrates into short-chain fatty acids, with CO2 and H2 as byproducts. (1) Organisms like Methanobrevibacter smithii convert hydrogen into methane, maintaining low levels of hydrogen in the intestines. (2) S. Typhimurium can utilize hydrogen as an energy source to increase invasion into intestinal epithelial cells. M. smithii sequesters hydrogen and competes with S. Typhimurium, limiting its virulence. (3) S. Typhimurium, and to a limited degree S. Typhi, causes intestinal inflammation and ROS production by immune cells. Anti-oxidant encoding-genes in M. smithii are upregulated in response to oxidative stress during S. Typhi human infection, potentially detoxifying and balancing the redox environment and limiting S. Typhi disease progression.
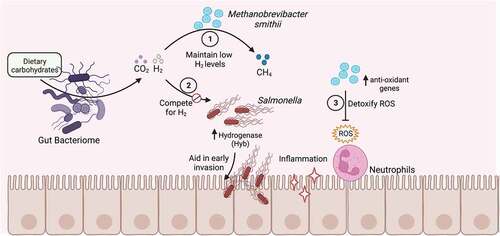
Figure 4. Direct interactions of gut fungi with enteric bacterial pathogens.
Fungi within the gut microbiome interact and influence enteric bacterial pathogens by different mechanisms, potentially impacting their virulence. (1) Fungi can directly bind bacteria and reduce their binding to the gut epithelium. (2) Secreted molecules including quorum sensing molecules, such as farnesol and ethanol, can affect bacterial biofilm formation, virulence, and growth. (3) Fungi and bacteria can form mixed bacterial biofilms, which protect the bacterial pathogen against antibacterial drugs and immune cells. (4) Salmonella Typhimurium can utilize siderophores produced by fungi to acquire iron in iron-limited environments. (5) Helicobacter pylori can enter and grow inside the vacuole of Candida. (6) C. albicans provides a favorable environment for the growth of the anaerobic bacterium Clostridium difficile by utilizing oxygen in its vicinity.
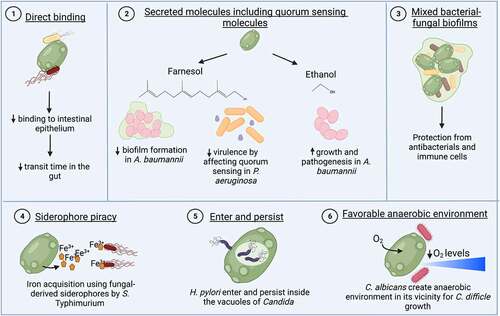
Figure 5. Indirect effects of the gut mycobiome on enteric bacterial infections.
(1) Gut fungi are involved in modulating the composition and colonization of commensal gut bacteria, which can in turn affect the colonization resistance to bacterial pathogens. (2) The fungal cell wall is composed of a chitin skeleton linked to a network of β-1,3-glucans and β-1,6-glucans, anchoring proteins and mannans toward the cell surface. Fungal cell wall components act as pathogen-associated molecular patterns (PAMPs) and are recognized by the immune cells via different pathogen recognition receptors (PRRs). The activation of immune cells such as macrophages by these PAMPs induce the release of IL-22 from Th17 cells. IL-22 plays a key role in maintaining gut barrier function against enteric pathogens. (3) Dendritic cells associated with Peyer’s patches recognize C. albicans and signal to promote the accumulation of regulatory T cells. The accumulation is dependent on the induction of 2,3-indoleamine dioxygenase (IDO) by dendritic cells. (4) C. tropicalis induces the migration of gut RALDH+ dendritic cells to the peripheral lymph nodes and helps in the maturation of the lymph nodes. These suggested mechanisms help to maintain mucosal immunity.
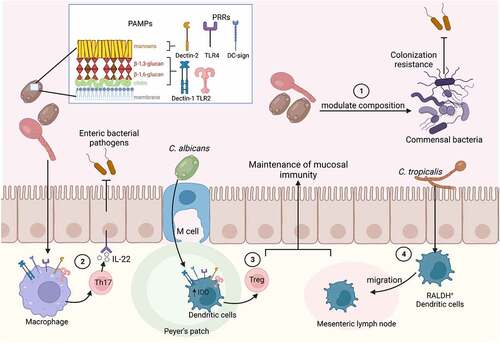
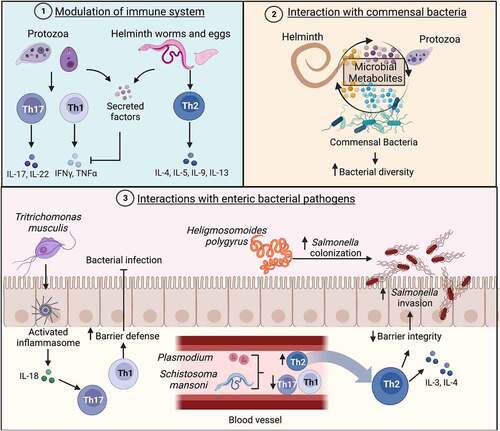
Figure 6. Effects of the gut parasitome on enteric bacterial infections.
(1) Protozoa parasites induce Th1 and Th17 responses while helminths induce Th2 response. Secreted parasitic effectors inhibit pro-inflammatory cytokines. (2) Members of the gut parasitome interact with commensal bacteria and secrete metabolites that may serve to increase bacterial diversity. (3) The parasite Tritrichomonas musculis activates the epithelial cell inflammasome to induce production of IL-18, which leads to a Th1 and Th17 response that increases barrier defense and limits bacterial infection. The presence of Heligmosomoides polygyrus enhances S. Typhimurium colonization, although the mechanism is currently unknown. Blood-associated parasites, like Plasmodium species and Schistosoma mansoni, activate a Th2 response, which leads to IL-3 and IL-4 production, disrupting the epithelial barrier in the gut. S. Typhimurium takes advantage of this environment and shows increased epithelial cell invasion.