
ABSTRACT
Epimedium is the largest and one of the most important herbaceous genera in Berberidiaceae, especially for section Diphyllon, which has a diversification center in central China. Chromosome numbers and karyomorphology of 24 accessions were examined, representing 18 Epimedium species, five of which were counted here for the first time. A chromosome number of 2n = 2x = 12 was observed for all taxa and all karyotypes were Stebbins’ 2A, except for E. acuminatum, E. brevicornu, E. ilicifolium and one accession of E. leptorrhizum, which were 3A. Their asymmetry index (AI) values vary from 1.71 to 4.56. Intercalary satellites are commonly found on the first or second pair. More than two individuals were sampled for E. franchetii, E. leptorrhizum, E. stellulatum, and E. wushanense and all had consistent and stable cytological traits with respect to their karyotype formula, Stebbins’ asymmetry, and AI values, except for E. leptorrhizum. The significance and limits of the cytological results for Epimedium are also briefly discussed.
1. Introduction
Epimedium Linnaeus (Citation1753), a morphologically diverse genus, is the largest perennial herbaceous genus in Berberidaceae, with more than 50 species widely disjunct between the Mediterranean region and eastern Asia (Stearn Citation2002; Ying Citation2002; Zhang et al. Citation2006). With more than 40 species, China is the most important center of diversity for Epimedium (Ying Citation2002; He and Xu Citation2003; Guo et al. Citation2007, Citation2008; Zhang and Li Citation2009; He et al. Citation2010; Sheng and Tian Citation2011; Ying et al. Citation2011; Zhang et al. Citation2014a, Citation2015, Citation2016; Wei et al. Citation2017). Moreover, all of these species are endemic to China except for E. koreanum Nakai, which is also found in Korea (Ying et al. Citation2011). Epimedium is well-known as a traditional Chinese medicine that is reported to be effective in strengthening the kidneys, curing rheumatism, and treating osteoporosis, hypertension, and coronary heart disease, as well as strengthening immunity and preventing dementia (Guo and Xiao Citation1999; Sun et al. Citation2005). Icariin flavonoids from Epimedium have been shown to inhibit the growth of cancer cells (Zhang et al. Citation2002; Xie and Sun Citation2006).
Recent molecular phylogenetic studies have suggested that the genus is differentiated into five groups largely corresponding to the taxonomical classification of two subgenera and four sections (Stearn Citation2002; Sun et al. Citation2005; Zhang et al. Citation2014b). Stearn (Citation2002) treated all Chinese species as subgenus Epimedium section Diphyllon, which is further split into four series based on flower morphology. Although amplified fragment length polymorphism (AFLP) evidence suggests that five well-supported clades are recognized in Epimedium sect. Diphyllon, the backbone relationship of the section remains unclear (Zhang et al. Citation2014b). Species delimitation in Epimedium has also long been in debate due to diverse morphological traits, especially for the Chinese species of Epimedium sect. Diphyllon, which has wide variations in petal type, form, the relative size of the inner sepals and petals, and flower dimensions (Ying Citation2002; Zhang et al. Citation2006, Citation2015; Smet et al. Citation2012).
Cytological data, such as chromosome numbers and karyotypes, are very important in understanding plant evolution and diversification (Stebbins Citation1950, Citation1971; Hong Citation1990; Stace Citation2000), which are significant characters in attempting to deduce phylogenetic relationships in angiosperms (Raven Citation1975). To date, more than 40 Epimedium taxa have reported chromosome numbers and/or karyotypes (e.g. Tanaka and Takahashi Citation1981; Takahashi Citation1989; Sheng and Chen Citation2007, Citation2008; Zhang et al. Citation2008; Sheng et al. Citation2010; Yan et al. Citation2016). All examined taxa have somatic chromosome numbers of 2n = 2x = 12, except for E. yingjiangense with a somatic chromosome number of 2n = 4x = 24 (Sheng and Tian Citation2011). Furthermore, the cyto-morphology of all previously examined taxa closely resemble one another with respect to size and shape of chromosomes, with most characterized as relatively primitive 2A and a few as 1A or 3A (Zhang et al. Citation2008; Sheng et al. Citation2010; Yan et al. Citation2016). A secondary constriction has commonly been found on the proximal side of the short arm of chromosome pair I or II for most species (Sheng and Chen Citation2008; Sheng et al. Citation2010).
With the report of many recently discovered Chinese species, the interspecific relationships of Epimedium sect. Diphyllon have become more complex, further confusing the delimitation and systematic arrangement of the species in this genus. The recent interest in understanding the development of Epimedium has prompted the need for in-depth taxonomic and phylogenetic studies of the genus, especially for the highly diverse Chinese taxa in Epimedium sect. Diphyllon. Chromosome numbers and karyotypes have been already reported for a large number of Chinese species from sect. Diphyllon (Sheng and Chen Citation2007, Citation2008; Zhang et al. Citation2008; Sheng et al. Citation2010; Yan et al. Citation2016); however, more than one fifth of the Chinese species are lacking chromosome counts. It is necessary to further accumulate cytological data for Epimedium spp. from China. In the present study, we report chromosome numbers and karyotypes from Epimedium sect. Diphyllon species from China and then evaluate possible infrageneric and intergeneric relationships based on our results and previous data.
2. Materials and methods
Cytological examinations were performed on somatic metaphases of root-tips using the squash technique. A total of 24 populations, representing 18 species of Epimedium, were collected between 2016 and 2017 in central China and cultivated in a nursery at Jishou University, Hunan, China. Voucher specimens were deposited in the Herbarium of Jishou University (JIU). Fresh root tips c.1 cm long were cut, pretreated in 1/100 colchicine solution at 22–24°C in the dark for 4 h and then fixed with Carnoy’s fluid (1:3 glacial acetic acid/absolute alcohol) at approximately 4°C for 40 min. The root tips were then placed in 70% ethanol for 10 min and rinsed in distilled water twice for 20 min. Prior to staining, the root tips were hydrolyzed in 1:1 1 N HCl to 45% acetic acid at 60°C for 30 s, stained with 1% aceto-orcein overnight, and squashed for chromosome observations. The preparations obtained were examined and photographed using a Leica DM1000 microscope (Leica Microsystems, Wetzlar, Germany).
Metaphasic plates or idiograms were analyzed with KaryoType software (http://mnh.scu.edu.cn/soft/blog/KaryoType/index.html) and lengths of individual chromosomes were measured. Using karyotype data for each species, chromosome pairs were arranged in order of decreasing length, and then the absolute lengths of the long and short arms of each chromosome pair were measured. The data were then pooled according to the considered taxonomic groups. In order to provide the general phenotypic appearance of karyotypes, a mean haploid idiogram (see also Peruzzi et al. Citation2009) was built for each sample.
The designation of the centromeric position as median (m), submedian (sm), or subterminal (st) followed methods described by Levan et al. (Citation2010), and the karyotype asymmetries were estimated according to Stebbins (Citation1971) and Paszko (Citation2006). The following parameters were calculated for the numerical characteristics of the karyotypes: (1) shortest (SC) and longest (LC) chromosome length; (2) ratio of the longest to shortest chromosome (LC/SC); (3) mean long arm length (p); (4) mean short arm length (q); (5) mean total chromosome length (CL); (6) asymmetry index (AI) = CVCL × CVCI/100 (Paszko Citation2006); (7) Stebbins’ type (KA); and (8) karyotypic formula ().
3. Results
Detailed chromosome parameters of studied taxa are shown in , and karyomorphology and idiograms are illustrated in .
Table 1. Sample information and karyotypic characteristics of 18 Epimedium species examined in this study.
3.1. Epimedium acuminatum Franchet
The karyotype formula of E. acuminatum collected from the Emei Mountains in Sichuan is 2n = 2x = 12 = 6m (2sat) + 6sm. The first, third, and fifth chromosome pairs are metacentric and the others are submetacentric as observed at mitotic metaphase. Intercalary satellites were observed on the first pair of chromosomes. The total haploid length of the chromosomes is 22.00 μm with single chromosome lengths ranging from 2.93 to 4.36 μm. The Stebbins’ asymmetry is categorized as 3A and the AI = 2.26 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
Figures 1-24. Metaphase chromosomes of the 18 examined species in Epimedium. 1, E. acuminatum; 2, E. baojingense; 3, E. brevicornu; 4–6, E. franchetii; 7, E. hunanense; 8, E. ilicifolium; 9–11, E. leptorrhizum; 12, E. lishihchenii; 13, E. membranaceum; 14, E. mikinorii; 15, E. parvifolium; 16, E. pseudowushanense; 17, E. sagittatum; 18, E. shennongjiaense; 19, 20, E. stellulatum; 21, E. tianmenshanensis; 22, 23, E. wushanense; 24, E. zhushanense. Scale bar = 5 μm.
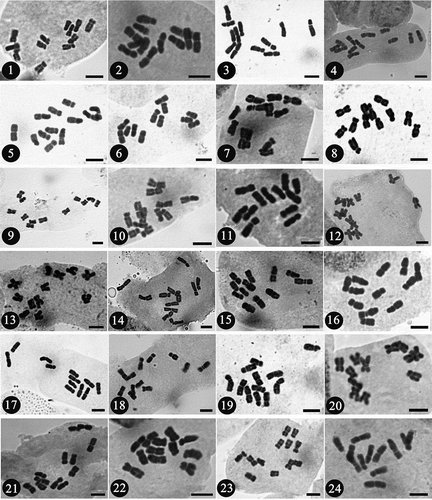
Figure 25-48. Idiograms of the 18 species of Epimedium examined in this study. 25, E. acuminatum; 26, E. baojingense; 27, E. brevicornu; 28–30, E. franchetii; 31, E. hunanense; 32, E. ilicifolium; 33–35, E. leptorrhizum; 36, E. lishihchenii; 37, E. membranaceum; 38, E. mikinorii; 39, E. parvifolium; 40, E. pseudowushanense; 41, E. sagittatum; 42, E. shennongjiaense; 43, 44, E. stellulatum; 45, E. tianmenshanensis; 46, 47, E. wushanense; 48, E. zhushanense. Scale bar = 2 μm.

3.2. Epimedium baojingense Q. L. Chen & B. M. Yang
The somatic chromosome number of E. baojingense is 2n = 2x = 12 = 6m (2sat) + 6sm, with satellites observed in the interstitial region of the first pair of chromosomes. The first, second, and fifth chromosome pairs are metacentric and the others are submetacentric as observed at mitotic metaphase. The individual chromosome lengths range between 4.06 and 5.38 μm, with a total haploid length of 27.98 μm. The Stebbins’ asymmetry category is 2A and the AI = 2.14 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
3.3. Epimedium brevicornu Maximowicz
Epimedium brevicornu collected from Shanxi has a somatic karyotype formula of 2n = 2x = 12 = 4m (2sat) + 8sm. Its first and third chromosome pairs are metacentric and the others are submetacentric as observed at mitotic metaphase. Satellites were observed in the interstitial region of the second pair of chromosomes. The chromosome lengths range between 4.03 and 5.84 μm and the total haploid length of the chromosome is 29.65 μm. The Stebbins’ asymmetry is categorized as 3A and the AI = 2.37 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
3.4. Epimedium franchetii Stearn
Three samples of E. franchetii were included in this study, and two types of karyotype formulas and different AI values were observed; the same Stebbins’ asymmetry category was found (). The first sample, collected from Wufeng, Hubei, has a somatic chromosome number of 2n = 2x = 12 = 6m (2sat) + 6sm. In this sample, the first, second, and third chromosome pairs are metacentric and the others are submetacentric as observed at mitotic metaphase. Intercalary satellites were observed on the first pair of chromosomes. The chromosomes lengths range between 3.9 and 5.95 μm and the total haploid length of the chromosome is 27.41 μm. The Stebbins’ asymmetry category is 2A and the AI = 2.68 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
The second sample, also collected from Wufeng, Hubei, has a somatic karyotype formula of 2n = 2x = 12 = 6m (2sat) + 6sm. At mitotic metaphase, the first, second and third chromosome pairs are metacentric and three pairs of submetacentric chromosomes were observed. Intercalary satellites were observed on the second pair of chromosomes. The chromosome lengths range between 3.26 and 4.61 μm. The total haploid length of the chromosome is 23.86 μm. The Stebbins’ asymmetry is 2A and the AI = 2.28 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
The third sample was collected from Shennongjia, Hubei. The somatic karyotype formula is 2n = 2x = 12 = 6m (2sat) + 6sm and its first, second, and third chromosome pairs are metacentric and the others are submetacentric as observed in mitotic metaphase. Satellites were observed in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.28 and 4.77 μm and the total haploid length of the chromosome is 24.26 μm. The Stebbins’ asymmetry is classified to 2A and the AI = 2.57 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
3.5. Epimedium hunanense (Handel-Mazzetti) Handel-Mazzetti
The chromosome number and karyotype is first reported for E. hunanense. The mitotic metaphase chromosomes and idiogram for this species are given in and , respectively. Its somatic karyotype formula is 2n = 2x = 12 = 6m (2sat) + 4sm + 2st. At mitotic metaphase, the first, the second and third chromosome pairs are metacentric, the fourth, and fifth pairs are submetacentric, and the sixth pair is subteleocentric. Satellites were observed in the interstitial region of the first pair of chromosomes. Chromosome lengths range between 3.73 and 5.87 μm. The total haploid length of the chromosome is 27.44 μm. The Stebbins’ asymmetry is categorized as 2A and the AI = 4.56 ().
3.6. Epimedium ilicifolium Stearn
The somatic karyotype formula of E. ilicifolium is 2n = 2x = 12 = 6m (2sat) + 6sm. At mitotic metaphase, the first, third and fourth chromosome pairs are metacentric, the second, fifth, and sixth pairs are submetacentric. Satellites were found in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.24 and 4.95 μm and the total haploid length of the chromosome is 25.08 μm. The Stebbins’ asymmetry category is 3A and the AI = 2.88 (). The mitotic metaphase chromosomes and idiogram are presented in and , respectively.
3.7. Epimedium leptorrhizum Stearn
Three samples of E. leptorrhizum were investigated, showing two types of karyotype formulas, different AI values, and different Stebbins’ asymmetry categories. The first sample, collected from Longshan, Hunan, has a chromosome number of 2n = 2x = 12 = 6m (2sat) + 6sm. At mitotic metaphase, the first, second, and third chromosome pairs are metacentric and the others are submetacentric. Satellites were observed in the interstitial region of the second pair of chromosomes. The chromosome lengths range between 2.88 and 4.04 μm with a total haploid length of 29.25 μm. The Stebbins’ asymmetry category is classified as 2A and the AI = 2.19 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
Epimedium leptorrhizum was only recently reported in Zhejiang (Xia et al. Citation2011). The sample from Longquan, Zhejiang has a somatic karyotype formula of 2n = 2x = 12 = 6m (2sat) + 6sm. At mitotic metaphase, the first, second, and third chromosome pairs are metacentric and the others are submetacentric. Satellites were observed in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 2.87 and 4.57 μm with a total haploid length of 20.19 μm. The Stebbins’ asymmetry category is 3A and the AI = 3.31 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
The third sample, collected from Enshi, Hubei, has a somatic karyotype formula of 2n = 2x = 12 = 4m + 8sm (2sat) with two pairs of metacentric and four pairs of submetacentric chromosomes. Satellites were observed in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 4 and 5.76 μm and the total haploid length is 22.69 μm. The Stebbins’ asymmetry is 2A and the AI = 1.94 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
3.8. Epimedium lishihchenii Stearn
The mitotic metaphase chromosomes and idiogram of E. lishihchenii are given in and , respectively. Its somatic karyotype formula is 2n = 2x = 12 = 6m (2sat) + 6sm. The first, second, and fourth chromosome pairs are metacentric and the others are submetacentric as observed at mitotic metaphase. The second pair of chromosomes bear satellites at an intercalary position. The chromosome lengths range from 2.98 to 4.5 μm and the total haploid length of the chromosome is 21.56 μm. The Stebbins’ asymmetry is 2A and the AI = 2.10 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
3.9. Epimedium membranaceum K. Meyer
The chromosome number and karyotype of E. membranaceum was counted for the first time with samples collected from Dujiangyan, Sichuan. The karyotype formula is 2n = 2x = 12 = 6m (2sat) + 6sm. The first, second, and third chromosome pairs are metacentric and the others are submetacentric as observed at mitotic metaphase. Satellites are located in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.21 and 4.54 μm and the total haploid length of the chromosome is 23.25 μm. The Stebbins’ asymmetry is categorized as 2A and the AI = 2.23 (). The mitotic metaphase chromosomes and idiogram are presented in and , respectively.
3.10. Epimedium mikinorii Stearn
Epimedium mikinorii, collected from Enshi, Hubei, has a somatic karyotype formula of 2n = 2x = 12 = 6m (2sat) + 6sm. At mitotic metaphase, the first, second, and third chromosome pairs are metacentric and the others are submetacentric. Satellites are located in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.45 and 5.02 μm, with a total haploid length of 25.91 μm. The Stebbins’ asymmetry category is 2A and the AI = 2.40 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
3.11. Epimedium parvifolium S. Z. He & T. L. Zhang
The somatic karyotype formula of E. parvifolium is 2n = 2x = 12 = 6m (2sat) + 6sm. The first, second, and fourth chromosome pairs are metacentric and the others are submetacentric as observed at mitotic metaphase. Satellites were observed in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.13 and 5.53 μm. The total haploid length of the chromosome is 24.29 μm. The Stebbins’ asymmetry is 2A and the AI = 2.76 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
3.12. Epimedium pseudowushanense B. L. Guo
The chromosome number and karyotype formula of E. pseudowushanense from Longli, Guizhou is 2n = 2x = 12 = 6m (2sat) + 6sm. At mitotic metaphase, the first, second, and third chromosome pairs are metacentric and the others are submetacentric. Satellites were observed in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.81 and 5.68 μm. The total haploid length of the chromosome is 27.63 μm. The Stebbins’ asymmetry is categorized as 2A and the AI = 1.71 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
3.13. Epimedium sagittatum (Siebold & Zuccarini) Maximowicz
The somatic karyotype formula of this sample from Youyang, Chongqing is reported with 2n = 2x = 12 = 6m (2sat) + 6sm. The first, second, and third chromosome pairs are metacentric and the others are submetacentric as observed at mitotic metaphase. Satellites are located in the interstitial region of the first pair of chromosomes. The chromosome lengths range from 3.78 to 5.65 μm with the total haploid length of 27.23 μm. The Stebbins’ asymmetry is 2A and the AI = 2.66 (). The mitotic metaphase chromosomes and idiogram are presented in and , respectively.
3.14. Epimedium shennongjiaense Y. J. Zhang & J. Q. Li
Epimedium shennongjiaense, a new species reported by Zhang and Li (Citation2009), is endemic to Shennongjia, Hubei Province. The chromosome number is counted here for the first time with the somatic karyotype formula of 2n = 2x = 12 = 4m + 8sm (2sat). The second and third pairs of chromosomes are metacentric and the others are submetacentric as observed at mitotic metaphase. Satellites were found in the interstitial region of the first pair of chromosomes. The chromosome lengths range from 3.72 to 5.14 μm and the total haploid length is measured as 25.40 μm. The Stebbins’ asymmetry category is 2A and the AI = 1.85 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
3.15. Epimedium stellulatum Stearn
Two samples of E. stellulatum were included here, having different karyotype formulas and AI values, but the same Stebbins’ asymmetry. The somatic karyotype formula of the sample from Wudang Mountain, Hubei is 2n = 2x = 12 = 8m (2sat) + 4sm. It has four pairs of metacentric and two pairs of submetacentric chromosomes found at mitotic metaphase. Satellites were observed in the interstitial region of the first pair of chromosomes. Its chromosome lengths range between 3.6 and 5.56 μm with the total haploid length of 26.72 μm. The Stebbins’ asymmetry is 2A and the AI = 1.90 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
The second sample was collected from Luonan, which is a new record in Shaanxi. The somatic karyotype formula is 2n = 2x = 12 = 6m (2sat) + 6sm. Three pairs of metacentric (1–3) chromosomes were observed and the other chromosomes were submetacentric as investigated at mitotic metaphase. Satellites were located in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.45 and 4.99 μm and the total haploid length is 25.15 μm. The Stebbins’ asymmetry is categorized as 2A and the AI = 2.89 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
3.16. Epimedium tianmenshanensis T. Deng, D. G. Zhang & H. Sun
Epimedium tianmenshanensis is a recently described species (Zhang et al. Citation2015), endemic to the Tianmen Mountain in western Hunan Province. Its somatic karyotype formula was first examined here: 2n = 2x = 12 = 6m (2sat) + 6sm. The first, second, and third pairs of chromosomes are metacentric and the others are submetacentric at mitotic metaphase. Satellites were observed in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.47 and 5.42 μm and the total haploid length of the chromosome is 26.29 μm. The Stebbins’ asymmetry is 2A category and the AI = 3.71 (). The mitotic metaphase chromosomes and idiogram are presented in and , respectively.
3.17. Epimedium wushanense T. S. Ying
Two counts were reported for this species with different karyotype formulas and AI values, but the same Stebbins’ asymmetry. The first sample was collected from a low elevation (500 m) area of Xiagu Town, Shennongjia National Nature Reserve, Hubei. The somatic karyotype formula is 2n = 2x = 12 = 8m (2sat) + 4sm. At mitotic metaphase, the first, second, fourth, and fifth pairs of chromosomes are metacentric and the others are submetacentric. Satellites were observed in the interstitial region of the first pair of chromosomes. The chromosome lengths range from 3.83 to 5.64 μm. The total haploid length of the chromosome is 27.12 μm. The asymmetry category is Stebbins’ 2A and the AI = 1.90 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
The second sample was collected from Shennongjia National Nature Reserve in Hubei. Its somatic karyotype formula is 2n = 2x = 12 = 4m + 8sm (2sat). The second and third chromosome pairs are metacentric and the other chromosomes are submetacentric as found at mitotic metaphase. Satellites were observed in the interstitial region of the first pair of chromosomes. The chromosome lengths range from 2.97 to 4.25 μm and the total haploid length is 21.80 μm. The Stebbins’ asymmetry is 2A and the AI = 2.22 (). The mitotic metaphase chromosomes and idiogram are shown in and , respectively.
3.18. Epimedium zhushanense K. F. Wu & S. X. Qian
Epimedium zhushanense is endemic to Hubei and its somatic chromosome number and karyotype formula is reported as 2n = 2x = 12 = 4m + 8sm (2sat). The second and third pairs of chromosomes are metacentric and the others are submetacentric as observed at mitotic metaphase. Satellites are located in the interstitial region of the first pair of chromosomes. The chromosome lengths range between 3.66 and 5.78 μm and its total haploid length is 29.09 μm. The Stebbins’ asymmetry is categorized as 2A and the AI = 2.76 (). The mitotic metaphase chromosomes and idiogram are given in and , respectively.
4. Discussion
This study reported chromosome numbers and karyotypes of 18 species of Epimedium from 24 populations (; –), all from sect. Diphyllon and all endemic to China. The cytological data confirmed that the chromosome numbers of E. acuminatum, E. baojingense, E. brevicornu, E. franchetii, E. ilicifolium, E. leptorrhizum, E. mikinorii, E. parvifolium, E. pseudowushanense, E. sagittatum, E. stellulatum, E. wushanense, and E. zhushanense are diploid (2n = 12) with the basic number of x = 6. Chromosome numbers of five species were reported for the first time (i.e. E. hunanense, E. lishihchenii, E. membranaceum, E. shennongjiaense, and E. tianmenshanensis) with 2n = 2x = 12 ().
Sheng et al. (Citation2010) reported the karyotype of Epimedium lishihchenii sampled from Tongren, Guizhou, with the somatic karyotype formula of 2n = 2x = 12 = 4m (2sat) + 8sm. However, their sample might be misidentified because E. lishihchenii is endemic to Jiangxi. Our material was collected from Lushan, Jiangxi with the chromosome number and karyotype correctly reported for the first time.
Combined with results reported in earlier studies (e.g. Tanaka and Takahashi Citation1981; Takahashi Citation1989; Sheng and Chen Citation2007, Citation2008; Sheng et al. Citation2010; Yan et al. Citation2016), cytological data has suggested that Epimedium has an evolutionary history of diploidy with 2n = 2x = 12, except for E. yingjiangense with 2n = 4x = 24 (Sheng and Chen Citation2007; Sheng and Tian Citation2011), but E. yingjiangense is also known to have 2n = 2x = 12 (Sheng and Chen Citation2007).
All Epimedium species examined in this study present a similar karyomorphology including only metacentric and submetacentric chromosomes, except for E. hunanense with its sixth chromosome pair subteleocentric with an arm ratio of 3.17 (). Previous studies also confirm similar karyotypes for Epimedium (Tanaka and Takahashi Citation1981; Takahashi Citation1989; Sheng and Chen Citation2007, Citation2008; Zhang et al. Citation2008; Sheng et al. Citation2010). In the present study, intercalary satellites were commonly found in the first pair in all species, except for E. leptorrhizum, E. lishihchenii, E. brevicornu, and E. franchetii, which had a secondary constriction in the second pair. The length of pairs 1 and 2 for the above-mentioned four species was similar. Combined with previous karyotype investigations on Epimedium (Tanaka and Takahashi Citation1981; Takahashi Citation1989; Sheng and Chen Citation2007, Citation2008; Zhang et al. Citation2008; Diao et al. Citation2011), we suggest that chromosome pair 2 of these four species was congenetic with pair 1 of the other species, which is similar to results found in Zhang et al. (Citation2008) and Sheng et al. (Citation2010).
Karyotype asymmetry indices have been widely used to make assumptions about the mechanisms of chromosomal evolution in plants (Paszko Citation2006). The Stebbins’ (Citation1971) classification is the most frequently used qualitative method for assessing symmetry conditions and describing the karyotypic relationships between different taxa. In this study, all of the investigated species display relatively primitive 2A asymmetry, except E. acuminatum, E. brevicornu, E. ilicifolium, and one accession of E. leptorrhizum, which showed 3A asymmetry (). Other previous studies have found a few species (E. baojingense, E. glandulosopilosum, E. leptorrhizum, E. pauciflorum, E. shuichengense, and E. yingjiangense with 1A asymmetry; E. dolichostemon with 3A asymmetry) with varying Stebbins’ classifications (Tanaka and Takahashi Citation1981; Takahashi Citation1989; Sheng and Chen Citation2007, Citation2008). As a result, the karyotypes of Epimedium sect. Diphyllon species investigated was highly uniform and primitive, as suggested by previous reports on Epimedium (Tanaka and Takahashi Citation1981; Takahashi Citation1989; Sheng and Chen Citation2007, Citation2008). Due to the uniform karyotype and the limited Stebbins (Citation1971) qualitative classification, it is difficult to infer the phylogenetic relationship within Epimedium solely based on karyotypic evidence.
Previous studies have suggested stable cytological results for species such as E. acuminatum, E. ecalcaratum, E. pubescens, and E. sagittatum (Sheng and Chen Citation2007, Citation2008; Zhang et al. Citation2008; Sheng et al. Citation2010; Diao et al. Citation2011). To further examine the chromosome variation within Epimedium species, more than two individuals were sampled for E. franchetii, E. leptorrhizum, E. stellulatum, and E. wushanense (). These species had consistent and stable cytological traits with respect to their karyotype formula, Stebbins’ asymmetry, and AI values, except for E. leptorrhizum. The two accessions of E. leptorrhizum from Hunan and Hubei have similar karyotypes and 2A asymmetry, but the third accession from Zhejiang had 3A type asymmetry and the AI = 3.31 (). The sample collected from Zhejiang is morphologically similar to the samples from Hunan and Hubei, but had relatively large flowers. Thus, further molecular phylogenetic studies are merited to test the status of the population of E. leptorrhizum from Zhejiang. Furthermore, this species has previously been reported with Stebbins 1A asymmetry (Sheng et al. Citation2010), which might indicate that chromosome variation is large within this species.
A recent molecular phylogeny of Epimedium conducted by Zhang et al. (Citation2014b) suggested that the two subgenera and four sections are monophyletic and Epimedium sect. Diphyllon was divided into five well-supported clades. This study also showed that five species (E. brevicornu, E. elongatum, E. pubescens, E. stellulatum, and E. sutchuenense) are either isolated or formed a polytomy located near the root of the section. Due to the uniform karyotypes and similar asymmetry in sect. Diphyllon, we found no distinct correlation between the karyotypic data and phylogenetic relationships among these Chinese species of Epimedium. Furthermore, we used a newer asymmetry index developed by Paszko (Citation2006), which has advantages of allowing a high degree of precision and sensitivity when assessing karyotype asymmetry. The AI values of the taxa included in this study range from 1.71 to 4.56, but no association was found with morphological characters or biogeographic distribution. As a result, it is difficult to discuss the phylogenetic evolution within Chinese Epimedium species based on the cytological results. Further evidence, such as fluorescent banding and physical mapping of 45S and 5S rDNA as shown in Sheng and Wang (Citation2010) and Diao et al. (Citation2011), provide promising advances in elucidating the evolutionary relationships among Epimedium species on the chromosomal structural level.
Acknowledgments
We thank Min Gao, Li-Ying Luo, and Zhen-Xin Xie for assistance in cytological lab works and Meng-Hua Zhang for her valuable comments on the early version of this manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Diao Y, Chen Q, Lin X, Liao C, Zhong LH, Wang YW. 2011. Karyotype characterization of Epimedium species (Berberidaceae) by fluorescent banding and physical mapping of 45S and 5S rDNA. Caryologia. 64:84–90.
- Guo BL, He SZ, Zhong GY, Xiao PG. 2007. Two new species of Epimedium (Berberidaceae) from China. Acta Phytotaxonomica Sinica. 45:813–821.
- Guo BL, Pei LK, Xiao PG. 2008. Further research on taxonomic significance of flavonoids in Epimedium (Berberidaceae). J Systematics Evol. 46:874–885.
- Guo BL, Xiao PG. 1999. The flavonoids in Epimedium L. and their taxonomic significance. Acta Phytotaxonomica Sinica. 37:228–243.
- He SZ, Wang YY, Guo BL, Xu WF. 2010. Epimedium pudingense (Berberidaceae), a new species from Guizhou, China. Annales Botanici Fennici. 47:226–228.
- He SZ, Xu WF. 2003. A new species of Epimedium (Berberidaceae) from Guizhou. Acta Botanica Yunnanica. 25:281–282.
- Hong DY. 1990. Plant cytotaxonomy. Beijing: Science Press.
- Levan A, Fredga K, Sandberg AA. 2010. Nomenclature for centromeric position on chromosomes. Hereditas. 52:201–220.
- Linnaeus C. 1753. Species plantarum. Stockholm: Laurentius Salvius, p. 117.
- Paszko B. 2006. A critical review and a new proposal of karyotype asymmetry indices. Plant Syst Evol. 258:39–48.
- Peruzzi L, Leitch IJ, Caparelli KF. 2009. Chromosome diversity and evolution in Liliaceae. Ann Bot. 103:459–475.
- Raven PH. 1975. The bases of angiosperm phylogeny: cytology. Ann Mo Bot Garden. 62:724–764.
- Sheng MY, Chen QF. 2007. Karyotype analysis of six Epimedium Species native to Guizhou,China. Guihaia. 27:440–443.
- Sheng MY, Chen QF. 2008. Giemsa C-banding analysis in eight species of genus Epimedium (Berberidaceae). J Wuhan Bot Res. 26:433–436.
- Sheng MY, Tian XJ. 2011. A new species of Epimedium (Berberidaceae) with 24 chromosomes from Guizhou, China. Novon J Bot Nomencl. 21:262–265.
- Sheng MY, Wang LJ. 2010. Chromosomal localization of 45S and 5S rDNA in 14 species and the implications for genome evolution of genus Epimedium. Plant Syst Evol. 290:65–73.
- Sheng MY, Wang LJ, Tian XJ. 2010. Karyomorphology of eighteen species of genus Epimedium (Berberidaceae) and its phylogenetic implications. Genet Resour Crop Evol. 57:1165–1176.
- Smet YD, Goetghebeur P, Wanke S, Asselman P, Samain MS. 2012. Additional evidence for recent divergence of Chinese Epimedium (Berberidaceae) derived from AFLP, chloroplast and nuclear data supplemented with characterisation of leaflet pubescence. Plant Ecol Evol. 145:121–126.
- Stace CA. 2000. Cytology and cytogenetics as a fundamental taxonomic resource for the 20th and 21st centuries. Taxon. 49:451–477.
- Stearn WT. 2002. The genus Epimedium and other herbaceous Berberidaceae. London: The Royal Botanic Gardens, Kew.
- Stebbins GL. 1950. Variation and evolution in plants. New York (NY): Columbia University Press.
- Stebbins GL. 1971. Chromosomal evolution in higher plants. London: Edward Arnold.
- Sun Y, Fung KP, Leung PC, Shaw PC. 2005. A phylogenetic analysis of Epimedium (Berberidaceae) based on nuclear ribosomal DNA sequences. Mol Phylogenet Evol. 35:287–291.
- Takahashi C. 1989. Karyomorphological studies on speciation of Epimedium and its allied Vancouveria with special reference to C-bands. Journal of Science of the Hiroshima. J Sci Hiroshima B, Division 2 (Botany). 22:159–269.
- Tanaka R, Takahashi C. 1981. Comparative karyotype analysis in Epimedium species by C-banding (1) E. sempervirens var. hypoglaucum and E. perralderianum. Jpn Bot. 56:17–24.
- Wei N, Zhang YJ, Zhun XU, Kamande EM, Ngumbau VM, Hu GW. 2017. Epimedium zhaotongense (Berberidaceae), a new species from Yunnan Province, China. Phytotaxa. 296:88.
- Xia GH, Yu L, Xie WY, Liu CH. 2011. Two new records of seed plants from Zhejiang, China. Journal of Zhejiang A & F University. 28(4):685–686.
- Xie JP, Sun WJ. 2006. Progress of chemical materials and pharmacy of genus Epimedium plants. Strait Pharm J. 18:17–20.
- Yan FL, He SZ, Xu WF, Li ZH, Pharmacy DO. 2016. Cytotaxonomy of seven species of Epimedium medicinal plants from Chinese. Guihaia. 36(9):1039–1045.
- Ying TS. 2002. Petal evolution and distribution patterns of Epimedium L. (Berberidaceae). Acta Phytotaxonomica Sinica. 40:481–489.
- Ying TS, Boufford DE, Brach AR. 2011. Berberidaceae. In: Wu ZY, Raven PH, editors Flora of China. Beijing: Science Press and Missouri Botanical Garden Press; p. 714–800.
- Zhang DG, Deng T, King C, Zhang JW, Nie ZL, Sun H. 2015. Epimedium tianmenshanensis (Berberidaceae), a new species from central China. Phytotaxa. 222:33.
- Zhang L, Wang Y, Mao HT. 2002. Study on the inhibition of telomerase activity and regulated mechanism in human cancer cell by Icarrin. Chin J Immunol. 18:191–196.
- Zhang ML, Kadereit HU, Joachim WK. 2006. Phylogeny and Biogeography of Epimedium/Vancouveria (Berberidaceae): western North American - East Asian Disjunctions, the Origin of European Mountain Plant Taxa, and East Asian Species Diversity. Syst Bot. 32:81–92.
- Zhang YJ, Dang HS, Li JQ, Wang Y. 2014a. The Epimedium wushanense (Berberidaceae) species complex, with one new species from Sichuan, China. Phytotaxa. 172:39–45.
- Zhang YJ, Dang HS, Meng AP, Li JQ, Li XD. 2008. Karyomorphology of Epimedium (Berberidaceae) and its phylogenetic implications. Caryologia. 61:283–293.
- Zhang YJ, Li JQ. 2009. A new species of Epimedium (Berberidaceae) from Hubei, China. Novon J Bot Nomencl. 19:567–569.
- Zhang YJ, Yang L, Chen J, Sun W, Wang Y. 2014b. Taxonomic and phylogenetic analysis of Epimedium L. based on amplified fragment length polymorphisms. Sci Hortic. 170:284–292.
- Zhang YJ, Zhang SJ, Dang HS, Zheng B, Li JQ, Wang Y. 2016. Epimedium xichangense (Berberidaceae), a new species from Sichuan, China. Phytotaxa. 3:263.