
ABSTRACT
Hamra National Park in the province of Dalarna in central Sweden was established in 1909 to protect a unique unlogged old-growth forest. In 1922, Henrik Hesselman set up two permanent plots in Hamra to be able to follow the development of the forest over time. In 2022, we re-inventoried the Scots pine-dominated plot and found that (1) the number of trees in the plot almost doubled from 593 trees in 1922 to 1013 trees in 2022, (2) Norway spruces are starting to take over the initially Scots pine-dominated plot, (3) the basal area of dead trees has increased by 286% and (4) the variability in annual Scots pine growth has increased over the studied time period. Given this strong deviation from the original state of the forest, we suggest restoring part of the forest to its early-1900s state while allowing the permanent plot to evolve naturally.
Introduction
In 1909, the first national parks were established in Sweden (Grundsten Citation1983) with the intention of saving “original” nature. The framework and the ideas for the set-up of larger protected areas were inspired by the establishment of national parks in the United States at the end of the nineteenth century (Grundsten Citation1983), but also with inspiration from European countries (Starbäck Citation1904). An important influencer at the time was a visiting German scientist, Professor Hugo Conwentz. He and others strongly advocated for the protection of important parts of the remaining “natural landscape” (Conwentz Citation1904; Starbäck Citation1904). An interesting argument was also that knowledge of the natural development of for example forests, were needed as “research stations” contrasting managed forests (Starbäck Citation1904). In 1905 Professor Einar Lönnberg brought up the idea of protecting an unlogged pristine forest (“urskog” in Swedish) as a future reference, and so that the development of this forest could be followed in the future (Lönnberg Citation1912). The suggestion was discussed at the Royal Academy of Sciences in Sweden and a formal decision to protect 20 hectares of Hamra National Park in the province in Dalarna was taken in 1909 (Lönnberg Citation1912).
A few years later, researchers started to document the unique Scots pine forest: In 1922, Professor Henrik Hesselman, a pioneer forest ecologist in Sweden, set up two sample plots in the forest and recorded all trees within these plots in a way so that each tree could be identified and its development followed over time. Such permanent plots in forests give precise spatial information of the state of the forest at a specific time, and they also provide unique opportunities for future researchers to analyse ecosystem change and dynamics if they are inventoried again after some time. The permanent plots in Hamra National Park are unique as they were established before the first commercial logging and modern forest management, making them by far the oldest in Scandinavia. Per Linder re-inventoried these plots in 1994 and evaluated the changes over the 72-year period (Linder Citation1998).
We revisited one of the two permanent plots in 2022 with the aim of investigating the changes in a natural Scots pine stand over a period of 100 years. Specifically, we wanted to answer the following questions:
How has the composition of species and the basal area of living and dead trees changed in the forest stand?
How has the annual growth of these Scots pine tree changed over the past century?
Based on our results, how could the forest be managed in the future?
Materials and methods
Study area and plot layout
The study area is located in Hamra National Park in central Sweden and the area was protected as an example of unlogged old-growth coniferous boreal forest in 1909 (Linder Citation1998). Initially, only 20 ha of the most pristine unlogged parts of the forest were protected (Linder Citation1998), but the national has since been enlarged and now spans 1383 ha including forests, mires and watercourses (Länsstyrelsen Gävleborg Citationno date).
In 1922, two permanent plots representing the two most common stand types in the park, open Scots pine (Pinus sylvestris L.) stands and mixed conifer stands dominated by Norway spruce (Picea abies (L.) H. Karst.), were established by Henrik Hesselman. He recorded the locations of all trees within the two 0.42 ha (60 m × 70 m) plots, tree status and species, and measured the diameters at breast height (Linder Citation1998).
Fieldwork in 2022
During the 100th anniversary year of these plots, we revisited the stand dominated by Scots pine and repeated the inventory done by Hesselman in 1922: We updated the status of each old tree (living, standing dead, or lying dead), added the locations of all new trees that have surpassed a height of 1.3 m since the last inventory done in 1994 (Linder Citation1998), and measured the diameters of all trees. In a subsection of the plot, we used a 5.15 mm increment corer to take one core sample each from 31 Scots pine trees that were already present in 1922, still alive and did not show any signs of damage or disease.
Data analyses and dendrochronology
For the analyses of the structural changes, we compared the data from 1922 to our data and evaluated the changes in species composition and basal area. For the basal area, we first calculated the cross-sectional area of each tree at breast height (1.3 m) using the measured diameter. These areas were then added up for the basal area of the 0.42 ha plot and finally scaled up to one hectare.
We prepared the cores according to standard dendrochronological methods (Schweingruber Citation1988), gluing them to wooden core mounts and then sanding the surface. After scanning them with a high-resolution scanner, we measured the ring-widths in CooRecorder and crossdated the samples with each other in CDendro (Larsson and Larsson Citation2022). Descriptive statistics for the tree-ring data and figures showing the mean chronology and distributions of age and diameter can be found in the supplementary material (Table 1; Figures 1–3).
Finally, we used the function bai.out() from the R package “dplR” (Bunn and Korpela Citation2020) to calculate the average basal area increment for each calendar year, i.e. the average two-dimensional annual growth of the sampled trees at breast height.
Results
Our results show that there have been drastic changes in both the species composition and stand structure over the 100-year time period (). The total number of trees in the 0.42 ha permanent plot has increased by 171% from 593 trees in 1922 to 1013 trees in 2022. While the number of Scots pine trees only increased by 38 trees from 439 in 1922 to 477 in 2022, there was a nearly 10-fold increase in the number of Norway spruce trees (from 49 in 1922 to 453 in 2022). Overall, young Norway spruces make up 91% of the trees that reached breast height after 1922.
Table 1. Changes in the number of living trees in the plot and the basal area of living and dead trees from 1922 to 2022.
While the number of trees is now very similar for both species, Scots pine trees still contribute significantly more to the basal areas of both the living trees (27.43 m2/ha Scots pine; 4.59 m2/ha Norway spruce) and dead trees (14.12 m2/ha Scots pine; 0.29 m2/ha Norway spruce). Overall, the basal area of living trees has increased by 156% over the 100-year period and the basal area of dead trees has increased by 286%. While Norway spruce only made up 3% of the basal area in 1922, this value has increased to 14% in 2022.
Contrary to the increase in the number of coniferous trees, the number of birch trees (Betula pubescens Ehrh.) has declined from 101 in 1922 to 57 in 2022. This translates to a 1042% increase in the basal area of dead birch from 1922 to 2022. However, the increase in the overall basal area of standing and lying deadwood from 5.12 m2/ha in 1922 to 14.63 m2/ha can be almost entirely attributed to the increase in dead Scots pine trees.
The number of non-birch deciduous trees – Goat willow (Salix caprea L.), Aspen (Populus tremula L.), Rowan (Sorbus aucuparia L.) and Grey alder (Alnus incana (L.) Moench) – has increased by 650% from only 4 trees in 1922 to 26 trees in 2022.
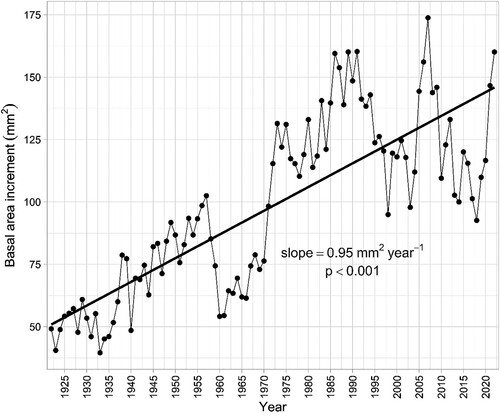
The dendrochronological analysis of a subset of the Scots pine trees that were already present in 1922 and still healthy in 2022 showed that these trees are growing much better now than 100 years ago (). On average, the basal area of each tree, i.e. the cross-sectional area at breast height, increased by 0.95 mm2/year from 49 mm2 in 1922 to 160 mm2 in 2022. From 1922 until 1957, the average basal area increment increased almost lineally before dropping slightly in the 1960s. More recently, the growth has varied more on a year-to-year basis than in the first half of the twentieth century.
Figure 1. Changes in annual average basal area increment of Scots pine trees in Hamra National Park from 1922 to 2022.
Discussion
We detected several trends in forest cover and species dynamics in the studied plot over the last 100 years (). First, the number of Norway spruces have increased dramatically, from a relative low number in 1922 (49) to very high numbers in 2022 (453). Norway spruces now contribute notably more to the forest cover than one hundred ago. Scots pine trees still contribute much more to the overall basal area than Norway spruce trees but this will obviously change as the spruces get bigger. As a consequence of the massive regeneration of Norway spruces, the forest has also become much denser over the past 100 years. These spruce thickets create different microclimatic conditions and alter the conditions for the flora and fauna inhibiting the stand. The amount of Scots pine deadwood has also increased dramatically.
The overall trend in increasing basal area increment () is similar to that of trees growing in a natural Scots pine forest in Tjeggelvas nature reserve in northern Sweden. Compared to this forest, annual Scots pine BAI is generally lower in Hamra National Park, which is particularly obvious when comparing the average growth during the drought year of 2018: While the trees in Tjeggelvas grew exceptionally well during that season (575 mm2), the mean growth in Hamra National Park was only 92 mm2 (Fassl Citation2023).
All these changes can be attributed to the lack of fire disturbance, which is the primary disturbance mechanism in natural boreal forest ecosystems (Zackrisson Citation1977), in the Scots pine stand during the studied time period. The most recent fire occurred in 1854 and absence of it since has favoured Norway spruce. This has ultimately caused a divergence from the original state of the forest in 1922 which was already noticed by Per Linder when he re-inventoried the plots in 1994 (Linder Citation1998). Wildfires very rarely affect forest reserves and national parks in Sweden, and prescribed burning is seldom practised in protected forests. This is primarily due to the high risks and costs associated with it.
Hedwall and Mikusiński (Citation2016) analysed National Forest Inventory (NFI) data to understand structural changes in forest reserves in Sweden over the last 50 years. They did not find any significant changes in tree species composition or intrusion of Norway spruce in Scots pine-dominated forests. However, it is important to note because of their different point of departure (1950s and 1960s; after the great transition of the boreal forest had already occurred), they also include more modern forest reserves that have a different history compared to the forest in Hamra National Park. In contrast to many forests reserves which were protected in the mid-twentieth century, the pre-industrial boreal forest in Hamra had a different structure with a sparse and open forest cover and was dominated by large and old trees (Lönnberg Citation1912; see also Östlund et al. Citation1997 Linder and Östlund Citation1998;). This further stresses the value and importance of long-term data, such as we have used in this study, to understand the consequences of active forest management vs non-intervention for the development of old-growth forest reserves.
Today, a non-intervention policy is guiding the management of Hamra National Park as well as many other protected forests in boreal Sweden. When the national park was established, it was stated that the motive for protection was “to hand down a primeval forest to posterity” (Linder Citation1998). However, non-intervention is also a form of management when it is applied to a dynamic ecosystem since it leads to a fundamental transition over time. Simply observing as the spruces gradually overtake will ultimately result in a forest that only partially resembles the original condition it was in for many centuries before its protection.
Different paths can be chosen for the future management of the old Scots pine stands in Hamra National Park. Non-intervention will eventually lead to a complete Norway spruce conversion of the forest, and a subsequent loss of the old Scots pines. Active management such as prescribed burning would mimic the historic disturbance regime, and would tentatively restore the forest to the state it was in one hundred years ago (Vanha-Majamaa et al. Citation2007). This method would be very challenging in the older parts of the forest due to the long time period since the last fire and subsequent accumulation of fuel in the ecosystem (Thomas and McAlpine Citation2010). To reduce the risk of crown fires, the majority of Norway spruce trees could be removed before a prescribed burning. An alternative to reduce competition and favour the growth of Scots pine would be to selectively harvest or kill Norway spruce trees. A local and traditional method to kill Norway spruce trees is to damage the cambial layer around the stem by using the blunt side of an axe (“taxning” in Swedish). This method can be used to selectively kill Norway spruce trees around the largest and oldest Scots pines in the forest reserve with the dual purpose of favouring Scots pines and creating Norway spruce deadwood.
The detailed registration of all trees in the sample plot in Hamra National Park in 1922 provides a unique window in time and allows us to understand the dynamics of an old-growth Scots pine forest and how the forest has deviated from its state when it was protected more than a century ago. In conclusion, we strongly advocate for deliberate yet subtle active management to restore the forest to a state that more closely resembles that of the early 1900s. In this specific case, we therefore suggest that the Scots pine-dominated permanent plot established by Hesselman in 1922 is left intact, while the forest surrounding is actively managed to restore and secure the ecological qualities of a unique Scots pine forest. This would also be in line with the initial motive for preservation, increase the ecological values of this particular forest, and allow for future investigations of the development of this forest.
Supplemental Material
Download MS Word (277.1 KB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
- Bunn A, Korpela M. 2020. An introduction to dplR. Seattle (WA): Washington University.
- Conwentz H. 1904. Om skydd åt det naturliga landskapet jämte dess växt- och djurvärld, särskildt i Sverige. Ymer. 24:17–42.
- Fassl M. 2023. Tree-growth and climate-growth relationships of Scots pine and downy birch in a natural forest in northern Sweden. Umeå: Sveriges lantbruksuniversitet.
- Grundsten C. 1983. Sveriges nationalparker. Solna: Statens naturvårdsverk.
- Hedwall PO, Mikusiński G. 2016. Sprucification in protected forests: myth or veracity? Clues from 60 yrs survey data. Appl Veg Sci. 19(3):371–380. doi:10.1111/avsc.12233.
- Länsstyrelsen Gävleborg. no date. Hamra. [accessed 2023 November 28]. https://www.lansstyrelsen.se/gavleborg/besoksmal/nationalparker/hamra.html.
- Larsson L, Larsson P. 2022. CDendro and CooRecorder (v.9.3.1). Saltsjöbaden: Cybis Elektronik. https://www.cybis.se.
- Linder P. 1998. Structural changes in two virgin boreal forest stands in central Sweden over 72 years. Scand J For Res. 13(1–4):451–461. doi:10.1080/02827589809383006.
- Linder P, Östlund L. 1998. Structural changes in three mid-boreal Swedish forest landscapes, 1885–1996. Biol Conserv. 85(1–2):9–19. doi:10.1016/S0006-3207(97)00168-7.
- Lönnberg E. 1912. Några ord om Hamra kronopark. Sveriges Natur. 1:72–75.
- Östlund L, Zackrisson O, Axelsson A-L. 1997. The history and transformation of a Scandinavian boreal forest landscape since the 19th century. Can J For Res. 27(8):1198–1206. doi:10.1139/x97-070.
- Schweingruber F. 1988. Tree rings: basics and applications of dendrochronology. Dordrecht: Kluwer.
- Starbäck K. 1904. Motion 1904:194 Andra kammaren: “Af herr K. Starbäck, om skrifvelse till Kungl. Maft angående skyddsåtgärder för vårt lands natur och naturminnesmärken” Motioner i Andra Kammaren, N:o 194, Sveriges Riksdag.
- Thomas PA, McAlpine RS. 2010. Fire in the forest. Cambridge: Cambridge University Press.
- Vanha-Majamaa I, Lilja S, Ryömä R, Kotiaho J, Laaka-Lindberg S, Lindberg H, Puttonen P, Tamminen P, Toivanen T, Kuuluvainen T. 2007. Rehabilitating boreal forest structure and species composition in Finland through logging, dead wood creation and fire: the EVO experiment. For Ecol Manag. 250(1–2):77–88. doi:10.1016/j.foreco.2007.03.012.
- Zackrisson O. 1977. Influence of forest fires on the North Swedish boreal forest. Oikos. 29:22–32. doi:10.2307/3543289.