
ABSTRACT
In recent years the high plateaus and escarpments of the north-eastern area of the Parataiko Range, Northland, Aotearoa – New Zealand have been recognised by taxonomists as a biodiversity hotspot with several new plant species unique to the whenua (land) above the Waima Valley. The wet and cool climate limits the growth of kauri (Agathis australis) but has allowed for unique species (e.g. Coprosma waima, Olearia crebra) to adapt to the steep-sided slopes and damp soils of the plateaus. Northland is also recognised as a biodiversity hotspot for the ground beetle genus Mecodema, with 15 described species endemic to this region, almost double that of an equivalent North Island entomological region. We describe, M. haakuturi sp. nov., which is restricted to the north-eastern arm of the Parataiko Range and is one of only a few Mecodema species found above an altitude of 500 m in Northland. We provide an updated key of Mecodema species from the Northland region, including the northern Auckland/Northland boundary species M. rusticulus and the widespread coastal M. antarcticum.
urn:lsid:zoobank.org:pub:CB67CF55-C486-4AFD-B0C1-5F7CAB86A419
Introduction
The entomological region of Northland (Crosby et al. Citation1998), North Island is regarded as a biodiversity hotspot due to its wide variety of unique habitats. The warm temperate climate and variety of substrates (e.g. volcanic, ultramafic, etc.) have produced a range of forest types, such as gum land forest, coastal dune forest and kauri (Agathis australis)-broadleaf forest, which have remained intact through successive glaciations (McKelvey and Nicholls Citation1959; Boswijk Citation2005; Lorrey et al. Citation2018).
Northland forest ecosystems have developed over long geographical time to produce a rich diversity of locally endemic flora and fauna in many groups not present elsewhere in New Zealand. For example, there is an exceptionally high number of endemic species among some iconic groups (e.g. Coprosma waima, koromiko (Veronica), Olearia crebra, land snails, ground beetles, stick insects and reptiles) (Druce Citation1989; de Lange Citation1998; Heenan and Cameron Citation2002; de Lange et al. Citation2002; Spencer et al. Citation2006; Chapple et al. Citation2009; Buckley and Bradler Citation2010; Seldon et al. Citation2012; Buckley and Leschen Citation2013; Larochelle and Larievière Citation2016; Seldon and Buckley Citation2019). The geology, history of volcanism and inundation, glaciation, climatic and biological factors have produced a uniquely diverse landscape of ecosystems and niches unlike the rest of the North Island (Hayward Citation1987, Citation2017; Newnham et al. Citation2013).
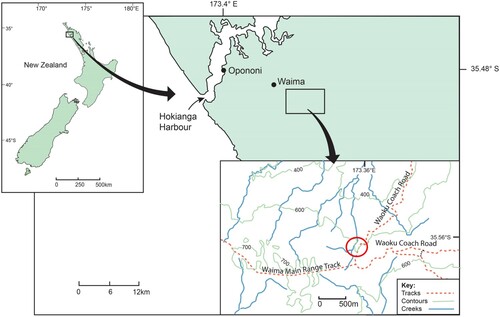
The inland north-eastern arm of the Parataiko Range (south of the Hokianga Harbour) is a unique area with steeply graded hillsides that make this mountain range relatively inaccessible (). The geology of this area is made up of underlying basalts and basaltic conglomerates of the Tangihua group, which are similar to most of the ranges in the western region (Hayward Citation1975, Citation1987, Citation2017). This north-eastern arm of the range, however, receives significantly more annual rainfall and is the coldest and highest (>700 m) part of Northland (Brock and Burns Citation2021). The northern area of the Mataraua Forest, within the Parataiko Range, has a series of plateaus (590–670 m) that are separated by steep escarpments (). The ground of the Mataraua Forest Plateau remains saturated throughout the year and, although it is part of the Northland temperate forest region (Boswijk Citation2005, Citation2010; Lorrey et al. Citation2018), the combined conditions of high altitude and high ground moisture produce a much colder microclimate than most of western Northland, limiting the growth of Agathis australis (kauri) (Brock and Burns Citation2021).
Figure 1. The approximate locality on the Waoku Coach Road Track (red circle), Parataiko Range, Northland, Aotearoa – New Zealand where the two specimens of Mecodema haakuturi sp. nov. were collected by the late John Ward, Canterbury Museum, Christchurch.
Mecodema species are found in almost all the Northland habitats (including dune forest systems, gum lands, kauri-podocarp-broadleaf, coastal forest) and, in recent revisions, 15 regionally endemic species () have been described (Seldon and Leschen Citation2011; Seldon et al. Citation2012; Seldon and Buckley Citation2019). Although Mecodema species inhabit all kauri forests, specimens are more readily collected in areas where kauri is scarce or absent, especially along the higher diversity forest edges (Seldon, personal observation). In this study we describe a new species of Mecodema that is found in a highly specific area in the Parataiko Range.
Table 1. Described regionally endemic Mecodema species (in alphabetical order), the type locality and the ranges within the Northland (ND) region as per Crosby et al. (Citation1998), New Zealand.
Methods and materials
Two male specimens of a new Mecodema species were recognised from material collected by the late John Ward (Canterbury Museum) from Waoku Coach Road Track, 650 asl, (Mataraua Forest) (), who was in the wetter areas of Northland searching for caddisflies (Trichoptera) (Patrick Citation2016).
Male genitalia were extracted using forceps, then macerated in cold 10% KOH solution for 24 h and rinsed with water. Excess tissue and membranes were removed using forceps and micro-dissecting scissors to expose the parameres and aedeagus. Left and right parameres were removed by cutting them from the base of the aedeagal lobe (). After photographing the intact aedeagal lobe, the endophallus was everted using fine-pointed forceps and a hooked minuten pin set into a long matchstick. All parts of the genitalia were kept in 100% glycerol, placed in silicon genitalic tubes and pinned with the specimen. No female specimens were available.
Figure 2. Male genitalia (aedeagus) structures (detached) of Mecodema haakuturi: (AL) aedeagal lobe, length 4.6 mm; (LP) left paramere (detached from AL), length 3.65 mm; (RP) right paramere (detached from AL), length 3.4 mm (EP) endophallus, length 3.9 mm (everted from AL). The Y-sclerite contains the sclerotised central spicule and associated structures (Photographs by author: DSS) [CMNZ specimen 2007.163.10642].
![Figure 2. Male genitalia (aedeagus) structures (detached) of Mecodema haakuturi: (AL) aedeagal lobe, length 4.6 mm; (LP) left paramere (detached from AL), length 3.65 mm; (RP) right paramere (detached from AL), length 3.4 mm (EP) endophallus, length 3.9 mm (everted from AL). The Y-sclerite contains the sclerotised central spicule and associated structures (Photographs by author: DSS) [CMNZ specimen 2007.163.10642].](/cms/asset/a34b1b61-1ea3-4421-a28a-d394d37f9716/tnzz_a_2334022_f0002_oc.jpg)
All photographs of the specimens for the habitus and genitalia illustrations were taken using a Visionary Digital Passport II System with a MP. E 65 mm lens, image series were processed using auto-stacker software Helicon Focus 5.3. The habitus and genitalia line drawings were prepared by Vivian Ward (School of Biological Sciences, University of Auckland) with the aid of Adobe Illustrator (Adobe Systems, California, USA). The habitus line drawing does not include the antenna or complete legs, as these characters are coded at genus level and are not used in species descriptions, except the width and shape of the protibia. Illustrations of the male genitalia structures are included and the left paramere (LP) and the right paramere (RP) have been drawn after being removed from the base of the penis lobe (PL) ().
The dorsal body length is measured from labrum to apical end of the elytra and the width at the widest point of both the pronotum and the elytra.
Results
This new species was discovered among unidentified North Island Mecodema specimens borrowed from the Canterbury Museum at the end of DS’s Doctorate research. The specimens are from an area of western Northland that is rugged and has steep-sided terrain, that is difficult to access, but rewarding as several new plant species and subspecies have recently been described from this area (personal communication, de Lange; +citations).
Taxonomy
Mecodema haakuturi sp. nov.
, .
Diagnosis
Distinguishable from other North Island Mecodema species by having: (1) the vertexal groove absent, except a few wrinkles sparsely distributed laterally; (2) asetose punctures of elytral striae star-shaped in the apical third (); and (3) regarding the apical shape of the aedeagal lobe, the form of the left paramere is distinctive with long setae extending along ⅘ of the ventral edge (, LP, AL).
Figure 3. Mecodema haakuturi sp. nov. plate: A, dorsal habitus; B, habitus illustration; C, ventral habitus. The scale bar under the habitus is the width across the widest point of the elytra of the specimen used for this illustration. Illustration by Vivian Ward, School of Biological Sciences, University of Auckland (Photographs by author: DSS) [CMNZ specimen 2007.163.10642].
![Figure 3. Mecodema haakuturi sp. nov. plate: A, dorsal habitus; B, habitus illustration; C, ventral habitus. The scale bar under the habitus is the width across the widest point of the elytra of the specimen used for this illustration. Illustration by Vivian Ward, School of Biological Sciences, University of Auckland (Photographs by author: DSS) [CMNZ specimen 2007.163.10642].](/cms/asset/2f19be62-bbf2-4b23-a4ed-6aeb3fada705/tnzz_a_2334022_f0003_oc.jpg)
Figure 4. Ventral view of a Mecodema specimen showing specific morphological structures, excluding taxonomic structures indicated in detail figures, used in the species descriptions. VLF = ventrite lateral foveae; VSP = ventrite setose punctures; MTC = metacoxa; MTVP = metaventrite process (with carina); MSC = mesocoxa; PC = procoxa; PS = prosternum; G = gena; PES = proepisternum; MSE = mesepisternum; MTE = metepisternum; V1–V6 = ventrites 1–6 (ventrites 1–3 may be fused); M = midline (dashed line, not a taxonomic structure) [Reproduced from Seldon & Buckley (Citation2019) with permission from copyright holder, Zootaxa].
![Figure 4. Ventral view of a Mecodema specimen showing specific morphological structures, excluding taxonomic structures indicated in detail figures, used in the species descriptions. VLF = ventrite lateral foveae; VSP = ventrite setose punctures; MTC = metacoxa; MTVP = metaventrite process (with carina); MSC = mesocoxa; PC = procoxa; PS = prosternum; G = gena; PES = proepisternum; MSE = mesepisternum; MTE = metepisternum; V1–V6 = ventrites 1–6 (ventrites 1–3 may be fused); M = midline (dashed line, not a taxonomic structure) [Reproduced from Seldon & Buckley (Citation2019) with permission from copyright holder, Zootaxa].](/cms/asset/2c0076a2-15fa-4845-93ca-ec222a1d3872/tnzz_a_2334022_f0004_ob.jpg)
Description
Length 20.8–22.5 mm, pronotal width 6.13–6.27 mm, elytral width 7.38–7.59 mm. Colour of entire body glossy black.
Head
Narrow and convex. Vertex smooth (very faint isodiametric pattern laterally); vertexal groove absent, except a few wrinkles sparsely distributed laterally posteriad of the supraorbital puncture (A, B); small supraorbital puncture bearing three setae; supraorbital grooves absent; frons without microsculpture (A, B), depressions or punctures; frontoclypeal suture indistinct, tentorial pits small; anterior area of clypeus with a setose puncture each side bearing one seta, without microsculpture. Anterior/apical margin labrum rounded laterally, slightly emarginate with three setae regularly spaced either side of the midline. Mentum lobes rounded, median process very broad and short, slightly angled upward (15°), moderately indentate; mentum setae absent. Submentum sclerite constriction narrow, strongly folded with four setae, two each side laterally distributed. Stipes with two basal setae. Gula pits small, suture indistinct, gula slightly convex and smooth. Gena with rugose, transverse wrinkles forming an isodiametric pattern laterally.
Prothorax
Prothoracic carina narrow the entire length, crenulated with six to eight setae each side (A, B), extended beyond anterior angle; posterior lateral sinuation evidently carinate, angled outward; pronotum narrow and flattened, overall shape rounded; midline slightly defined, but contiguous with anterior medial triangular impression, disc without microsculpture, depressions or punctures; pronotal foveae deep and narrow; anterior edge distinctly curved and posterior edge curved. Prosternum concave with very fine transverse lines (C); proepisternum with scattered pitting. Procoxal setose punctures absent; protibia distally expanded and shovel-like.
Elytra
Broad and laterally convex; humeral angle evenly convex; basal margin moderately curved and gently sloped to base, interval 1 extended to margin; lateral carina narrow in anterior two thirds, broadened in posterior third (A, B), extended to humeral angle; humerus with two closely distributed anterior setose punctures; suture impressed; striae with small, regularly distributed, asetose punctures, obsolescent in middle, then star-shaped at apex (A, B); striae equally impressed (visible to naked eye) laterally (A, B); intervals 1–9 weakly convex (intervals 1–4 may appear flat); interval microsculpture present as transverse lines; 7th strial setal pattern with two setae in the anterior half, and four to six setae in the posterior half, setose punctures small.
Ventral surface
Mesepisternum with scattered pitting (C); setose punctures present on mesocoxae (1) and metacoxae (3). Abdominal ventrites 1–6 lineate laterally, ventrites 3–5 with one setose puncture each side of midline; ventrite six setae present: ♂ with two-to-three evenly-spaced setose punctures each side of the midline, close to the flattened apical edge; ventrites 4 and 5 may be foveate laterally. Anterior metaventrite process (C, ) a triangle with a broad carina apically, that narrows laterally.
Male genitalia
Apical portion of aedeagal lobe asymmetrically rounded, ventral edge forms a rounded point that curves symmetrically backward to meet the dorsal edge at the junction with the endophallus membrane (, AL); apical portion with slight deflection to right of vertical axis (ventral view); apex shape narrow the entire length; aedeagal shaft is equally narrow the entire length (AL), ventral edge straight (ventral view), overall length of the aedeagal lobe slightly curved ventrally (lateral view). Structures of the endophallus: lateral form of the apex of the central spicule rounded; dorsal form of the apex of the central spicule narrow to broadened apically (EP, Y-sclerite), deflected to a 60˚ angle (dorsal view); setal (scales) coverage of the apical plate is extensive (76–100%) (, EP, Y-sclerite); left setose flange is small; right setose flange is absent. Left paramere basal lobe narrow and rectangular (, LP), vertically twisted, with slight anterior hump forming a 45˚ angle to a short and narrow arm; narrow terminal lobe with thick tuft of long setae, arm curved inwardly; long setae along ⅘ of ventral edge; ventral edge of basal lobe curved ventrally toward arm. Right paramere narrow and triangular (RP), with very long setae extended along the entire length of the ventral edge; apical third moderately inwardly curved, paramere twisted vertically.
Female genitalia
No female specimens are known.
Comments
The male aedeagal structures of M. haakuturi are unique among Northland Mecodema species.
Distribution
New Zealand, North Island, Northland, north-eastern arm of Parataiko Range, Waoku Coach Road track (). There are no coordinates with the label data, so we have provided approximate coordinates (−35.56, 173.55) that are situated between the Waoku Tk and a stream.
Holotype
CMNZ male labelled. 3818 Waipoua, Waimatenui, Waoku Track, 650 m, Ward JB, 1.i.99 / HOLOTYPE Mecodema haakuturi n. sp. design. DS Seldon and D Pou 2023.
Paratype
1♂, 3818 Waipoua, Waimatenui, Waoku Track, 650 m, Ward JB, 1.i.99 / PARATYPE Mecodema haakuturi n. sp. design. DS Seldon and D Pou 2023 (CMNZ).
Etymology
Hākuturi are kaitiaki (guardians) of the ngahere (forest). This species dwells in the highest altitude forest of Northland, and therefore can observe all other forested areas within the region.
A key to Mecodema from Northland entomological region (Crosby et al. Citation1998), which is adapted from Seldon and Buckley (Citation2019). The key includes the northern-most Auckland region species (M. rusticulus), that is found in forest remnants north of around Wellsford, and the widespread coastal species M. antarcticum ().
1. Ventrites 3–5 with numerous setose punctures each side of midline; protibia greatly expanded distally and distinctly curved inward … M. antarcticum
- Ventrites 3–5 without setose punctures or with 1–3 setose punctures each side of midline; protibia expanded distally and without marked curve inward … 2
2(1). Vertexal groove indistinct or absent … 3
- Vertexal groove distinct, marked narrow groove, punctures and/or rugose … 4
3(2). Frons without microsculpture; submentum sclerite with 4 setae (2 each side) … M. haakuturi sp. n.
- Frons with a large, shallow depression each side and lateral grooves; submentum sclerite with 6–8 setae … M. ngaiatonga
4(2). Vertexal groove with punctures … 5
- Vertexal groove without punctures … 12
5(4). Prothoracic carina with 7–10 setae each side (asymmetrical) … 6
- Prothoracic carina with 11+ setae each side (asymmetrical) … 10
6(5). Prothoracic carina broad entire; elytral explanate area (carina) broad entire … M. teroroa
- Prothoracic carina narrow entire or broadened anteriorly; elytral explanate area (carina) narrow entire … 7
7(6). Metacoxa with 3 setose punctures; anterior metaventral process carina broadened anteriorly, narrowed laterally … M. ngaitahuhu
- Metacoxa with 2 setose punctures; anterior metaventral process carina broad or narrow entire length … 8
8(7). Frons medially convex with large shallow depressions laterally … M. rusticulus
- Frons flat, microsculpture and depressions absent … 9
9(8). Vertexal groove with punctures entire length; submentum sclerite broad … M. papake
- Vertexal groove with punctures laterally only; submentum sclerite narrow … M. teparawhau
10(5). Vertexal groove with punctures entire length; clypeus with lateral and medial punctures bearing 3 setae; prothoracic carina broad along entire length … M. tewhara
- Vertexal groove with punctures or rugose microsculpture medially or laterally only; clypeus with lateral punctures bearing 2 setae, medial puncture absent; prothoracic carina narrow along entire length … 11
11(10). Vertexal groove with punctures medially only; submentum sclerite narrow; anterior metaventrite process carina broad entire length … M. kipjac
- Vertexal groove with punctures and/or rugose microsculpture laterally only; submentum sclerite broad; anterior metaventrite process carina narrow entire length … M. kokoroiho
12(4). Vertexal groove with distinct V-shape medially … 13
- Vertexal groove without distinct V-shape medially … 14
13(12). Mentum process apex notched; mesecoxa without setose punctures; anterior metaventrite process carina absent (Three Kings Is. only) … M. regulus
- Mentum process apex distinctly indentate; mesecoxa with one setose puncture; anterior metaventrite process carina narrow entire length … M. tenaki
14(12). Vertexal groove medially narrow, laterally broad; anterior/apical margin labrum slightly emarginate; pronotal posterior medial impression present … 15
- Vertexal groove narrow entire length; anterior/apical margin labrum straight or curved outwardly; pronotum posterior medial impression absent … 16
15(14). Body broad, length 27–34 mm; mentum process short, broad and apex distinctly indentate … M. xylanthrax
- 15(14). Body narrow, length 20–26 mm; mentum process short, narrow and apex slightly indentate (Poor Knights Is. only) … M. ponaiti
16(14). Vertex with strong rugose microsculpture; frons with lateral grooves; prosternum microsculpture absent … M. parataiko
- Vertex without microsculpture; frons without lateral grooves; prosternum with transverse lines … 17
17(16). Mentum process narrow, long with apex notched; submentum sclerite with 6–8 setae; prothoracic carina smooth … M. kokoromatua
- Mentum process narrow, short with apex moderately indentate; submentum sclerite with 4 setae; prothoracic carina crenulated … M. manaia
Discussion
Northland was once thought to be depauperate of Mecodema species, with only one species described by the mid-1960s, M. regulus (Three Kings Islands), from the curvidens group (Britton Citation1949, Citation1964). The few larger specimens collected from Northland were placed within M. spiniferum (Waitakere Ranges, Auckland) and were considered part of an Auckland–Northland complex (Larochelle and Larievière Citation2001). Therefore, the majority of collectors focussed on the South Island, which was deemed to have a wider range of unique habitats than the North Island. However, recent studies have concluded that there are 16 regionally endemic Mecodema species () in Northland (Seldon and Leschen Citation2011; Seldon et al. Citation2012; Seldon and Buckley Citation2019) and that this high endemism matches that of other taxa, including stick insects, ground beetles, plants, etc. (de Lange et al. Citation2002; de Lange and Rolfe Citation2008; Buckley and Bradler Citation2010; Buckley et al. Citation2014; Larochelle and Larievière Citation2016).
Northland’s long geological history of uplift, inundation and volcanism (Hayward Citation2017) and the continuous forest cover through consecutive periods of glaciation has produced a wide variety of unique habitats that have led to a high number of locally endemic taxa. Mecodema haakuturi is an example of a species that is restricted to the highest areas of Northland, where kauri is not found, which is often indicative of Mecodema species in the Northland/Auckland/Coromandel regions. Even though many Mecodema species occur in kauri dominated forests of northern North Island, within these habitats they are more readily found in the parts of the forest where the presence of kauri is low or absent (Seldon and Buckley Citation2019). The north-eastern arm of the Parataiko Range is made up of a series of plateaus and steep escarpments that are surrounded by very steep-sloped hillsides. This topography, the high annual rainfall and the lower-than-average Northland mean temperatures (Brock and Burns Citation2021) have produced a wet habitat with several unique plant species (Riddell Citation1999; de Lange and Rolfe Citation2008).
Currently, M. haakuturi is only known from a locality that intersects the Waoku Coach Road Track at an elevation of 650 m asl (). This area is the edge of native forest and pine plantation, above the Waima Valley and within the Mataraua Forest. The shape of the genitalic structures (, LP, RP, AL) indicate that this species may have been isolated from the other Northland Mecodema for a relatively long time. Mecodema haakuturi exhibits a similar short and broad body-shape of some other curvidens group species, that are adapted to specific habitats, e.g. M. antarcticum (coastal dunes), M. moniliferum (braided-rivers). However, M. haakuturi lacks the increased setation covering the ventrites of the species adapted to abrasive habitats.
Almost all of the native forest habitats north of the Dargaville–Bream Head Line, close to latitude 36˚ (Heenan et al. Citation2022), have both a curvidens group species (small) and a spiniferum group species (large) inhabiting the same locality. Little is known about the interactions of these sympatric species, and a recent survey of Bream Head, Whangarei showed that the two species usually only have a narrow area of overlap (Seldon Citation2019). For example, M. manaia (curvidens group) is found in the remnant broadleaf forest (higher) and M. tewhara (spiniferum group) is found in the kanuka (Kunzea sp.)/manuka (Leptospermum sp.) regenerating forest on the lower slopes (Seldon Citation2019). Similarly, the two specimens of M. haakuturi were collected along with a specimen of M. teroroa, which is more likely to be found in the native forest edge alongside the pine plantation. In terms of determining rarity and conservation status for this species, a survey of the escarpments and surrounding areas along the Waoku Track would be required (permits have been acquired).
Acknowledgements
We would like to thank Cor Vink (Lincoln University), Johno Ridden and other staff at the Canterbury Museum for the loan of Mecodema specimens and their patience regarding the return of those loans. We would also thank the late John B. Ward (Canterbury Museum) for collecting the only two specimens on his trip through Northland while he was looking for Trichoptera. We would like to thank Peter de Lange (Unitec Te Pūkenga, Mt Albert, Auckland) for his initial discussions about the significance of the new plant species found in the same area, and that this new Mecodema species is not from the Waima Forest. We thank Vivian Ward, School of Biological Sciences for the habitus drawing and other illustrations. We thank the people of Ngāpuhi for discussion regarding naming and access to their rohe (tribal land). We thank the journal reviewers for their time, expertise, and comments.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Blanchard C. 1853. Voyage au pole sud et dans l'Océanie sur les Corvettes l'Astrolabe et la Zélée; exécuté par ordre du roi pendant les années 1837–1838–1839–1840, sous le commandement de MJ Dumont-D'Urville, capitaine de vaisseau; publié par ordre du gouvernement, sous la direction supérieure de M. Jacquinot, capitaine de vaisseau, commandant de la Zélée. Zoologie par MM. Hombron et Jacquinot. Tome quatrième.
- Boswijk G. 2005. A history of kauri. Australia and New Zealand Forest Histories. Kingston, Australia: Araucarian Forests, Australian Forest History Society. pp. 19–26.
- Boswijk G. 2010. Remembering kauri on the ‘Kauri Coast’. New Zealand Geographer. 66:124–137. doi:10.1111/j.1745-7939.2010.01178.x
- Britton EB. 1949. The Carabidae (Coleoptera) of New Zealand. Part III. A revision of the tribe Broscini. Transactions of the Royal Society of New Zealand. 77:533–581.
- Britton EB. 1964. New Carabidae (Coleoptera) from Three Kings Islands, New Zealand. New Zealand Journal of Science. 7:521–527.
- Brock JMR, Burns BR. 2021. Patterns of woody plant epiphytism on tree ferns in New Zealand. New Zealand Journal of Ecology. 45:3433. https://www.jstor.org/stable/27041263
- Buckley TR, Bradler S. 2010. Tepakiphasma ngatikuri, a new genus and species of stick insect (Phasmatodea) from the Far North of New Zealand. New Zealand Entomologist. 33:118–126. doi:10.1080/00779962.2010.9722200
- Buckley TR, Leschen RA. 2013. Comparative phylogenetic analysis reveals long-term isolation of lineages on the Three Kings Islands, New Zealand. Biological Journal of the Linnean Society. 108:361–377. doi:10.1111/j.1095-8312.2012.02009.x
- Buckley TR, Myers SS, Bradler S. 2014. Revision of the stick insect genus Clitarchus Stål (Phasmatodea: Phasmatidae): new synonymies and two new species from northern New Zealand. Zootaxa. 3900:451–482. doi:10.11646/zootaxa.3900.4.1
- Chapple DG, Ritchie PA, Daugherty CH. 2009. Origin, diversification, and systematics of the New Zealand skink fauna (Reptilia: Scincidae). Molecular Phylogenetics and Evolution. 52:470–487. doi:10.1016/j.ympev.2009.03.021
- Crosby TK, Dugdale JS, Watt JC. 1998. Area codes for recording specimen localities in the New Zealand subregion. New Zealand Journal of Zoology. 25(2):175–183. doi:10.1080/03014223.1998.9518148
- de Lange PJ. 1998. Hebe perbella (Scrophulariaceae)—a new and threatened species from western Northland, North Island, New Zealand. New Zealand Journal of Botany. 36:399–406. doi:10.1080/0028825X.1998.9512578
- de Lange PJ, Gardner RO, Riddell KA. 2002. Ackama nubicola (Cunoniaceae) a new species from Western Northland, North Island, New Zealand. New Zealand Journal of Botany. 40(4):525–534. doi:10.1080/0028825X.2002.9512813
- de Lange PJ, Rolfe JR. 2008. Hebe saxicola (Plantaginaceae)—a new threatened species from western Northland, North Island, New Zealand. New Zealand Journal of Botany. 46:531–545. doi:10.1080/00288250809509783
- Druce AP. 1989. Coprosma waima (Rubiaceae)—a new species from northern New Zealand. New Zealand Journal of Botany. 27:119–128. doi:10.1080/0028825X.1989.10410150
- Hayward BW. 1975. Waipoua Basalt and the geology of Maunganui Bluff. Tane. 21:39–48.
- Hayward BW. 1987. Identity of igneous bodies beneath the continental shelf of west Northland, New Zealand. New Zealand Journal of Geology and Geophysics. 30:93–99. doi:10.1080/00288306.1987.10422197
- Hayward BW. 2017. Out of the ocean into the fire: history in the rocks, fossils and landforms of Auckland, Northland and Coromandel. Geoscience Society of New Zealand. 1–336.
- Heenan PB, Cameron EK. 2002. A new species of Olearia (Asteraceae) from Waima Forest, Northland, New Zealand. New Zealand Journal of Botany. 40:535–542. doi:10.1080/0028825X.2002.9512814
- Heenan PB, McGlone MS, Mitchell CM, Cheeseman DF, Houliston GJ. 2022. Genetic variation reveals broad-scale biogeographic patterns and challenges species’ classification in the Kunzea ericoides (kānuka; Myrtaceae) complex from New Zealand. New Zealand Journal of Botany. 60:2–26. doi:10.1080/0028825X.2021.1903946
- Larochelle A, Larievière M-C. 2001. Carabidae (Insecta: Coleoptera) catalogue. Fauna of New Zealand 43. Lincoln, New Zealand: Manaaki Whenua Press.
- Larochelle A, Larievière M-C. 2016. Taxonomic Supplement (2001–2015) to the Catalogue of New Zealand Carabidae (Insecta: Coleoptera). Insecta Mundi. 0502:1–53. http://digitalcommons.unl.edu/insectamundi/1010
- Lorrey AM, Boswijk G, Hogg A, Palmer JG, Turney CS, Fowler AM, Ogden J, Woolley JM. 2018. The scientific value and potential of New Zealand swamp kauri. Quaternary Science Review. 183:124–139. doi:10.1016/j.quascirev.2017.12.019
- McKelvey PJ, Nicholls JL. 1959. The indigenous forest types of North Auckland. New Zealand Journal of Forestry. 8:29–45.
- Newnham R, McGlone M, Moar N, Wilmshurst J, Vandergoes M. 2013. The vegetation cover of New Zealand at the last glacial maximum. Quaternary Science Reviews. 74:202–214. doi:10.1016/j.quascirev.2012.08.022
- Patrick BH. 2016. John Brigham Ward PhD, 10 January 1928–5 April 2016. New Zealand Entomologist. 39:147–149. doi:10.1080/00779962.2016.1210995
- Riddell K. 1999. Endemic plants of the Waima Range western Northland. Auckland Botanical Society Journal. 54:4–5.
- Seldon DS. 2019. A revision of the genus Mecodema Blanchard 1853 (Coleoptera: Carabidae: Broscini) from the North Island, New Zealand, and biogeographical interpretations of the subtribe Nothobroscina. PhD Thesis in Biological Sciences. University of Auckland.
- Seldon DS, Buckley TR. 2019. The genus Mecodema Blanchard 1853 (Coleoptera: Carabidae: Broscini) from the North Island, New Zealand. Zootaxa. 4598:1–148.
- Seldon DS, Leschen RAB. 2011. Revision of the Mecodema curvidens species group (Coleoptera: Carabidae: Broscini). Zootaxa. 2829:1–45. doi:10.11646/zootaxa.2829.1.1
- Seldon DS, Leschen RAB, Liebherr JK. 2012. A new species of Mecodema (Carabidae: Broscini) from Northland, New Zealand, with notes on a newly observed structure within the female genitalia. New Zealand Entomologist. 35:39–50. doi:10.1080/00779962.2012.649707
- Spencer HG, Brook FJ, Kennedy M. 2006. Phylogeography of kauri snails and their allies from Northland, New Zealand (Mollusca: Gastropoda: Rhytididae: Paryphantinae). Molecular Phylogenetics and Evolution. 38:835–842. doi:10.1016/j.ympev.2005.10.015