
ABSTRACT
Renowned for its leaf-spice attributes, kaffir lime has economic value that is worthy of intensifying amidst environmental threats, such as drought stress. This study aimed to analyze the morpho-physiological traits of kaffir lime in response to defoliation and drought stress. Kaffir lime was subjected to drought, defoliation, and a combination of both arranged in a randomised complete block design. Concerning morphology, defoliated lime produced a new flush that grows like a crown, while drought-treated plants displayed a bent pose with withered, edge-rolled, and drooping leaves. Higher leaf drop incidences were also noticed in drought-stressed plants. A combination of defoliation and drought stress produced severe flushing disruption by reducing 61% flush number, 77% flush fresh weight, and 74% flush dry weight, compared to undefoliated plants in normal irrigated conditions. Regardless of drought stress conditions, leaf production did not show significant differences among all defoliated plants. Defoliated plants maintained their photosynthetic rate and water use efficiency, leading to the absence of bent flush and rolling leaves as morphological markers of drought-treated plants. It is implied that defoliation can be a practical strategy for adapting kaffir lime in a drought environment.
Introduction
Global conditions are currently experiencing climate change phenomena that potentially endanger the agricultural sector (Sovacool et al. Citation2021; Bibi and Rahman Citation2023). Agricultural production should be performed in changing climatic variables, especially temperature, and rainfall intensity (Pope et al. Citation2022). Warmer global temperature has been noticed recently since there has been an increase of about 0.9°C from the nineteenth century (Eftekhari Citation2022). Moreover, the temperature in 2011–2022 was 1.1°C higher than that in the late nineteenth century, placing this finding as the warmest condition in the last 100,000 years (IPCC Citation2022). That change results in the incidence of extreme events such as floods, and drought (IPCC Citation2014; Bibi and Rahman Citation2023). Developing countries, especially in Asia and Sub-Saharan Africa, are the most vulnerable to being negatively impacted (IPCC Citation2014).
A developing country such as Indonesia is the place of origin of kaffir lime (Araujo et al. Citation2003), a unique citrus species with aromatic and bifoliate leaves (Budiarto et al. Citation2021a; Budiarto et al. Citation2023a) used for essential oils (Budiarto and Sholikin Citation2022) and food spice (Budiarto et al. Citation2019a) in numerous Asian cuisines (Setiyoningrum et al. Citation2018). Hence, the leaf became the main economic part (Efendi and Budiarto Citation2022) and is marketed in fresh, frozen, and dried forms (Wongpornchai Citation2012). The production and supply of kaffir lime leaf is also potentially hampered by climate change.
Climate change led to the incidence of prolonged hot and dry seasons, and ultimately resulted in drought stress problems (Ziogas et al. Citation2021). Drought stress is the most well-known limiting factor for citrus productivity and quality worldwide (Sato Citation2015). Previous studies have reported the effect of drought stress on citrus, i.e. alteration of gibberellin dan abscisic acid status (Mahouachi et al. Citation2005), antioxidants production for ROS scavenging (Hussain et al. Citation2018), photosynthetic rate reduction (Arjona-López et al. Citation2023), and eventually fruit yield drop (Zhang et al. Citation2018). Drought tolerance may differ among citrus species, as previously reported by Romero-Trigueros et al. (Citation2021) that grapefruit (Citrus paradisi) has greater tolerance than mandarin (Citrus clementina). However, drought-related effects have not been widely studied in kaffir lime, compared to other Citrus species.
To overcome drought stress, an agronomic solution such as leaf removal/pruning is potentially applied. The leaf removal practice successfully improved the fig (Ficus carica) plant tolerance to drought stress (Jafari et al. Citation2017; Zare Citation2021). Severe pruning applied under drought conditions impedes leaf chlorosis incidence (Zare et al. Citation2021), and helps to maintain a greater stomatal conductance (Maurin and DesRochers Citation2013). Moreover, pruned grapes can balance their berry composition and development (Vitis vinifera) under drought conditions (Berhe Citation2022). The agronomic practice of defoliation has been adopted in kaffir lime to harvest its leaves yield (Budiarto et al. Citation2019b; Budiarto et al. Citation2023a) however, its effect to alleviate drought-negative problems has not been reported yet. Here, the present work tests the hypotheses that drought and defoliation would affect the morpho-physiological properties of kaffir lime and that defoliation can be alleviated the negative effect of drought stress. This study provides a valuable reference for the agronomic solution to address drought issues in kaffir lime plantations.
Materials and methods
Kaffir lime material
This study used kaffir lime seedlings that originated from grafting, combining kaffir lime var. Puri Agrihorti on the Citrus limonia rootstock. Selected seedlings possessed several characteristics, i.e. plant height (±100 cm), leaf number (±40 leaves), normal growth, bifoliate leaf characteristics, pest- and disease-free. Eight months after grafting, seedlings grew in the polybag with a diameter of 30 cm, containing growing media in the form of a well mixture of clay soil, goat manure, and rice husk. The seedling was raised under organic culture by monthly application of liquid fermented goat manure.
Seedlings were arranged in the Randomised Completely Block Design inside the greenhouse at the Bale Tatanen Padjadjaran (−6.9316851, 107.775863, 718 m above sea level), from July to August 2023. The use of greenhouse is aimed to eliminate the bias effect of rainfall so that the only water source is through irrigation. The experiment employed two factors, i.e. defoliation and drought stress. The first factor of defoliation was comprised of two levels, i.e. defoliated and undefoliated seedlings. Defoliation was performed manually by eliminating all leaves from the seedling, while undefoliated seedlings had 40 leaves attached to their canopy. The second factor of drought stress was also comprised of two levels, i.e. normal and drought stress. The stress was created by the regulation of watering once in two weeks, while normal seedlings were exposed by irrigation once in two days. All combination treatments were replicated five times. During the experimental period, seedlings were raised by manual weeding and pest inspection once a week.
Effect of defoliation and drought stress
Concerning the effect of defoliation and drought stress, all morpho-physiological observations and measurements were carried out at 4 weeks after treatment (WATs). The morphological appearance of treated kaffir lime was observed and documented in figures. Morphological responses in terms of leaf drop occurrence were measured only in the unpruned kaffir lime seedlings due to the absence of old leaves in all defoliated seedlings. The drop leaves were calculated by comparing the leaf number at pre- and post-drought stress. The old leaves were counted based on the number of retained leaves at post-drought stress. The weight of old leaves was measured on a fresh and dry basis. Flush production of kaffir lime seedlings was measured in the form of a flush number (flush per plant), flush dry weight (g), flush fresh weight (g), and flush length (cm). The proportion of leaves and branches in newly emerged flush was counted and displayed as a chart. Individual leaf length was measured manually with a ruler, while leaf weight and leaf area were calculated, following an earlier study by Budiarto et al. (Citation2021b). Leaf production in response to defoliation and drought stress was also measured regarding leaf number (per plant), leaves dry weight (g), and leaves fresh weight (g). The leaves water content was the gap between fresh and dry weight and then was expressed in %.
Physiological traits of kaffir lime were measured by using the Li-6400XT instrument on a sunny day at 09.00 am. Intercellular CO2 (μmol CO2 mol−1), leaf temperature (°C), transpiration rate (mmol H2O m−2 s−1), photosynthetic rate (μmol CO2 m−2 s−1) and stomatal conductance (mol H2O m−2 s−1) were directly produced by that instrument, while intrinsic water use efficiency/WUE (μmol CO2 mmol H2O−1) was counted by dividing the rate of photosynthesis by the rate of transpiration, following previous study (Roccuzzo et al. Citation2014; Budiarto et al. Citation2023b). All data were collected and subjected to analysis of variance and then continued to the Duncan multiple range test at α 0.05. All statistical data were processed in RStudio version 2023.06.2 + 561.
Results
Different morphological appearance due to defoliation and drought stress
The morphological appearance of kaffir lime plants differs due to defoliation and drought stress. Defoliation is prepared by removing 40 leaves on the initial days, leaving only a single main stem with some newly emerged flush at the apical parts, irrespective of drought treatment (A, 1B). All defoliated seedlings display a similar flush appearance under drought-stressed and normal conditions (A, 2B). Without defoliation (C, 1D), drought-stressed kaffir lime plants exhibit a bent pose, while normal irrigated plants grow upright. In more detail, the bent flush of drought-stressed plants comprised withered, edge-rolled, and drooping leaves, while normal irrigated flush displays healthy and open-oriented leaves with towering growth patterns (C, 2D).
Figure 1. Morphological appearance of kaffir lime at 4 WATs subjected to various treatments: defoliation and drought stress (A); only defoliation (B); only drought stress (C); control (D). Note: The yellow line in the figure is equal to 100 cm.

Figure 2. Morphological appearance of kaffir lime flush at 4 WATs subjected to various treatments: defoliation and drought stress (A); only defoliation (B); only drought stress (C); control (D). Note: The yellow line in the figure is equal to 30 cm.

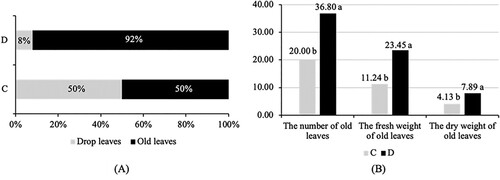
Higher leaf drop incidences in drought-threatened plants
This research proved that drought-solely increased leaf drop incidence by about 50%, compared to normally irrigated plants, which only experienced 8% leaf drop (A). 20 leaves still survive in the post-drought period, similar to the number of fallen leaves (B). In normally irrigated plants, only 3 leaves fall off due to leaf senescence. Consequently, old leaves’ fresh- and dry-weight variables of drought-stressed seedlings were also reduced by about 52% and 48% compared to normally irrigated seedlings, respectively (B).
Figure 3. The leaves drop occurrence leaving several old leaves on undefoliated kaffir lime seedlings during the post-drought stress period (4 WATs): the percentage of drop and old leaves (A) and the number and weight of old leaves (B). Note: Two compared seedlings experiencing leaf drop: drought stress (C); and control (D). Mean values above the bar inside the right chart followed by similar alphabet are not significantly different based on the Duncan Multiple Range Test at α = 0.05.
Flushing reduction by defoliation and drought condition
Flushing performance is an important component of leaf production, and it was significantly affected by defoliation and drought stress factors. Both factors significantly reduced flush number and flush weight; however, their effect was insignificant on flush length (). The highest significant reduction in flush number, flush fresh weight, and flush dry weight was found in the combination of defoliation and drought stress (treatment A) by about 61%, 77%, and 74% compared to control (treatment D), respectively. A significant reduction in flushing performance may be associated with a decline in vegetative growth ().
Table 1. Flushing characteristics of kaffir lime subjected to defoliation and drought condition.
Effect of defoliation is more dominant than drought on flush proportion and leaf size
In the newly emerged flush, the leaf proportion was more dominant than the branches ones; however, there was a tendency for defoliation to reduce the leaf proportion (). The higher leaf proportion in the newly emerged flush, as caused by the absence of defoliation, was supported by the individual leaf size data. Our finding in highlighted the lower individual leaf size of newly emerged flush in defoliated plants than undefoliated ones. The reduction of individual leaf area, individual leaf length, and individual leaf fresh weight were caused by defoliation by about 54%, 35%, and 56%, respectively () ().
Figure 4. The proportion of leaf and branch in newly emerged flush at 4 WATS. Note: Four compared treatments: defoliation and drought stress (A); only defoliation (B); only drought stress (C); control (D).
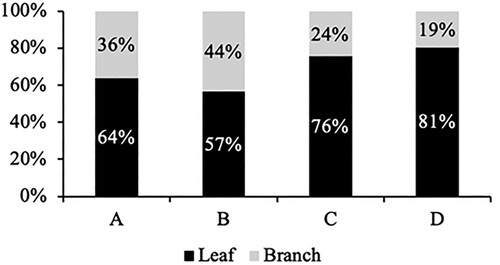
Table 2. Individual leaf size of kaffir lime subjected to defoliation and drought condition.
Drought stress drop leaf production of undefoliated plants, not in defoliated ones
Leaf production performances, as the most important variable in kaffir lime agribusiness, displayed a significant decline as caused by defoliation and drought condition. Leaf number, leaf weight, and water content were not significantly different among all defoliated plants, with or without drought stress exposure (). In contrast, undefoliated kaffir lime plants experienced a significant drop in leaf production if drought stress was applied. That reduction was reported to be 45% in leaf number, 57% in leaf fresh weight, 44% in leaf dry weight, and 15% in leaf water content ().
Table 3. Leaf production of kaffir lime subjected to defoliation and drought condition.
Table 4. Physiological properties of kaffir lime subjected to defoliation and drought condition.
Drought stress disrupt physiological properties of undefoliated plants, not in defoliated ones
depicted a significant reduction effect of drought condition (treatment C) on intrinsic WUE, stomatal conductance, and photosynthetic rate compared to control plants (treatment D) for about 34%, 35%, and 38%, respectively. The photosynthetic rate is not significantly reduced by defoliation, in contrast to the stomatal conductance variable that displays a significant reduction in defoliated and undefoliated plants was 12% and 35% compared to the control, respectively. Lower stomatal conductance is usually accompanied by lower intercellular CO2. However, opposite results were found in the present findings. Drought-subjected plants experienced a significantly higher intercellular CO2 and leaf temperature than normal irrigated plants, for about 28% and 11%. Additionally, the present finding also revealed a significant reduction of WUE in the drought-stressed, undefoliated plants by about 34% compared to the control. In contrast, the defoliated plant subjected to drought stress seemed to have a similar WUE to control.
Discussion
Drought and defoliation affect the morpho-physiology of kaffir lime
Concerning morphological properties, drought-stressed kaffir limes displayed a bent flush with edge-rolled, and drooping leaves (), similar to Hussain et al. (Citation2023) who reported leaf rolling and wilting in stressed sour oranges (Citrus aurantium). Within the flush, the upper leaves depicted higher leaf rolling intensities, implying a higher drought sensitivity than the middle and lower leaves () in agreement with Zhou et al. (Citation2021). Moreover, a high leaf drop incidence in drought-stressed plants (A) implied the alteration of hormonal status, i.e. an increase in abscisic acid content (Mahouachi et al. Citation2005) that triggers the abscission of the leaf stalk, (Rodriguez et al. Citation2019). Ultimately, drought stress declined leaf growth (Abobatta Citation2021; Shafqat et al. Citation2021), which would eventually cause a decrease in canopy size (Costa et al. Citation2021). Apart from drought, defoliation also formed a smaller leaf size, implying the reduction of plant source capacity due to photosynthate limitations after leaf removal; a similar case with Budiarto et al. (Citation2018) in mandarin and Yuan et al. (Citation2005) in oranges.
Concerning physiological responses, drought stress disrupted the photosynthesis rate of undefoliated limes, in agreement with Pena-Rojas et al. (Citation2004). Photosynthesis is suggested as a physiological marker of plant resistance to abiotic stress (Rahmat et al. Citation2023). The reduction of the photosynthesis rate could be caused by (i) a decline in stomatal conductance (Lawlor and Cornic Citation2002; Pena-Rojas et al. Citation2004), and (ii) leaf temperature-limiting factor, which was found only in undefoliated drought-stressed plants, as previously also reported by Lourkisti et al. (Citation2022). Taken together, this study proposes the morpho-physiological alteration of kaffir lime under leaf removal and drought stress conditions.
Defoliation alleviates the negative effect of drought stress on kaffir lime
In agreement with our hypothesis, defoliated kaffir limes were suggested to be more tolerant to drought () rather than undefoliated ones, as indicated by the absence of leaf rolling and wilting incidences. It was likely that defoliation helped to postpone the occurrence of drought symptoms (Namirembe et al. Citation2009). Pruned plants were supposed to have reduced water demand (Jackson et al. Citation2000; Lopez et al. Citation2012; Chen et al. Citation2016), as the consequence of the removal of certain canopy parts. The lowering water demand increases the WUE, which is proven in this study that the WUE of drought-exposed defoliated plants was as high as a control treatment with no drought conditions (). This work also showed no significant effect of defoliation on transpiration rate (p > 0.05) at the single leaf level; however, the declining leaf number in post-defoliated plants might suppress canopy area, and ultimately plant transpiration rate at the canopy level, as previously confirmed by numerous studies (Lopez et al. Citation2008; Jin et al. Citation2018; Devin et al. Citation2023). Pruned plant was reported to have a narrow xylem vessel, for lowering their susceptibility to xylem cavitation (Katul et al. Citation2003) and ultimately impede the rate of transpiration. In addition, pruned plants had greater stomatal control systems for optimising their net assimilation under stress conditions (Jin et al. Citation2018). This further supports the idea of defoliation as the feasible strategy to manage kaffir lime leaf production under drought-stress conditions.
Conclusion
The present study highlighted different morphological appearances of kaffir lime due to defoliation and drought stress. Defoliation removed a large portion of the existing canopy, leaving a new flush that grows like a crown. Normal irrigated plants grew upright, while drought-treated plants showed a bent pose comprised of withered, edge-rolled, and drooping leaves. Both defoliation and drought reduced flushing performance. However, the effect of defoliation is more dominant than drought on flush proportion and leaf size. Drought stress drop leaf production only in undefoliated plants, not in defoliated ones. Defoliated plants were able to maintain their photosynthetic rate and water use efficiency, leading to the absence of bent flush and rolling leaves as morphological markers of drought-treated plants. Pruning, especially defoliation, is strongly recommended, apart from harvesting activities, as a precautionary practice to plant kaffir lime in a drought environment.
Author contributions
RB, SM, and MAN were involved in the conception and design, MAR, and MHISJ in data analysis and interpretation; RB, and SA in the drafting of the paper, RB, DNS, AK, and SLS in revising it critically for intellectual content; RB in the final approval of the version to be published. All authors agree to be accountable for all aspects of the work.
Acknowledgments
The authors thank the team of Bale Tatanen Padjadjaran for the technical support during the fieldwork.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The data in the present finding are available from the corresponding author [RB] upon reasonable request.
Additional information
Funding
References
- Abobatta WF. 2021. Managing citrus orchards under climate change. MOJ Ecol Environ Sci. 6:43–44. doi:10.15406/mojes.2021.06.00212.
- Araujo EFD, Queiroz LPD, Machado MA. 2003. What is citrus? taxonomic implications from a study of cp-DNA evolution in the tribe Citreae (Rutaceae subfamily Aurantioideae). Organ Divers Evol. 3:55–62. doi:10.1078/1439-6092-00058.
- Arjona-López JM, Aparicio-Durán L, Gmitter FG Jr, Romero-Rodríguez E, Grosser JW, Hervalejo A, Arenas-Arenas FJ. 2023. Physiological influence of water stress conditions on novel hlb-tolerant citrus rootstocks. Agronomy. 13:63. doi:10.3390/agronomy13010063.
- Berhe DT. 2022. Post-veraison water stress and pruning level on merlot grapevine (Vitis vinifera L.): effects on berry development and composition. Int J Agronomy. 7307078:1–11. doi:10.1155/2022/7307078.
- Bibi F, Rahman A. 2023. An overview of climate change impacts on agriculture and their mitigation strategies. Agriculture. 13:1508. doi:10.3390/agriculture13081508.
- Budiarto R, Sholikin MM. 2022. Kaffir lime essential oil variation in the last fifty years: a meta-analysis of plant origins, plant parts and extraction methods. Horticulturae. 8:1132. doi:10.3390/horticulturae8121132.
- Budiarto R, Poerwanto R, Santosa E, Efendi D. 2018. Shoot manipulations improve flushing and flowering of mandarin citrus in Indonesia. J Appl Hortic. 20:112–118. doi:10.37855/jah.2018.v20i02.20.
- Budiarto R, Poerwanto R, Santosa E, Efendi D, Agusta A. 2019a. Production, post- harvest and marketing of kaffir lime (Citrus hystrix DC) in Tulungagung, Indonesia. J Trop Crop Sci. 6:138–143. doi:10.29244/jtcs.6.02.138-143.
- Budiarto R, Poerwanto R, Santosa E, Efendi D, Agusta A. 2019b. Agronomical and physiological characters of kaffir lime (Citrus 318 hystrix DC) seedling under artificial shading and pruning. Emirates J Food Agric. 31:222–230. doi:10.9755/ejfa.2019.v31.i3.1920.
- Budiarto R, Poerwanto R, Santosa E, Efendi D. 2021a. Morphological evaluation and determination keys of 21 citrus genotypes at seedling stage. Biodiversitas. 22:1570–1579. doi:10.13057/biodiv/d220364.
- Budiarto R, Poerwanto R, Santosa E, Efendi D, Agusta A. 2021b. Short communication: allometric model to estimate bifoliate leaf area and weight of kaffir lime (Citrus hystrix). Biodiversitas. 22:2815–2820. doi:10.13057/biodiv/d220545.
- Budiarto R, Mubarok S, Nanda MA, Nabiyyu M, Jaya MHIS. 2023a. The increase in kaffir lime leaf production due to gibberellin is diminished by pruning. Horticulturae. 9:1–11. doi:10.3390/horticulturae9091018.
- Budiarto R, Poerwanto R, Santosa E, Efendi D, Agusta A. 2023b. Agro-physiological traits of kaffir lime in response to pruning and nitrogen fertilizer under mild shading. Plants. 12:1–14. doi:10.3390/plants12051155.
- Chen D, Wang Y, Wang X, Nie Z, Gao Z, Zhang L. 2016. Effects of branch removal on water use of rain-fed jujube (Ziziphus jujuba mill.) plantations in Chinese semiarid loess plateau region. Agric Water Manag. 178:258–270. doi:10.1016/j.agwat.2016.10.010.
- Costa DP, Stuchi ES, Girardi EA, Moreira AS, Gesteira ADS, Filho MAC, Ledo CADS, da Silva ALV, de Leão HC, Passos OS, Filho WDSS. 2021. Less is more: A hard way to get potential dwarfing hybrid rootstocks for valencia sweet orange. Agriculture. 11:354. doi:10.3390/agriculture11040354.
- Devin SR, Prudencio AS, Mahdavi SME, Rubio M, Martínez-García PJ, Martínez-Gómez P. 2023. Orchard management and incorporation of biochemical and molecular strategies for improving drought tolerance in fruit tree crops. Plants. 12:773. doi:10.3390/plants12040773.
- Efendi D, Budiarto R. 2022. Benefit and challenges of using tropical fruits as ornamental trees for green city. Acta Hortic. 1334:369–378. doi:10.17660/ActaHortic.2022.1334.46.
- Eftekhari MS. 2022. Impacts of climate change on agriculture and horticulture. In: Bandh SA, editor. Climate change: the social and scientific construct. Cham, Switzerland: Springer International Publishing; p. 117–131.
- Hussain S, Sohail H, Noor I, Ahmad S, Ejaz S, Ali MA, Haider STA, Sohail M, Jaffer H, Ercisli S, et al. 2023. Physiological and biochemical determinants of drought tolerance in tetraploid vs diploid sour orange citrus rootstock. J Hortic Sci Biotechnol. 98:772–785. doi:10.1080/14620316.2023.2192230.
- Hussain S, Khalid MF, Saqib M, Ahmad S, Zafar W, Rao MJ, Morillon R, Anjum MA. 2018. Drought tolerance in citrus rootstocks is associated with better antioxidant defense mechanism. Acta Physiol Plant. 40:135, 1–10. doi:10.1007/s11738-018-2710-z.
- IPCC. 2022. Climate change 2021: Summary for All. Intergovernmental Panel on Climate Change, Geneva, Switzerland. https://www.ipcc.ch/report/ar6/wg1/downloads/outreach/IPCC_AR6_WGI_SummaryForAll.pdf.
- IPCC. 2014. Climate change 2014: impacts, adaptation, and vulnerability. contribution of working group II to the fifth assessment report of the intergovernmental panel on climate change. Cambridge, New York: Cambridge University Press.
- Jackson NA, Wallace JS, Ong CK. 2000. Tree pruning as a means of controlling water use in an agroforestry system in Kenya. For Ecol Manag. 126:133–148. doi:10.1016/S0378-1127(99)00096-1.
- Jafari M, Rahemi M, Zare H. 2017. Increasing the tolerance of fig tree to drought stress by trunk thinning. Acta Hortic. 1173. doi:10.17660/ActaHortic.2017.1173.32.
- Jin S, Wang Y, Shi L, Guo X, Zhang J. 2018. Effects of pruning and mulching measures on annual soil moisture, yield, and water use efficiency in jujube (Ziziphus jujube Mill.) plantations. Global Ecology and Conservation. 15:e00406. doi:10.1016/j.gecco.2018.e00406.
- Katul G, Leuning R, Oren R. 2003. Relationship between plant hydraulic and biochemical properties derived from a steady-state coupled water and carbon transport model. Plant Cell Environ. 26:339–350. doi:10.1046/j.1365-3040.2003.00965.x.
- Lawlor DW, Cornic G. 2002. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 25:275–294. doi:10.1046/j.0016-8025.2001.00814.x.
- Lopez G, Girona J, Marsal J. 2008. Drought in peach orchards: summer pruning vs. fruit thinning for water stress mitigation and long-term effects of water stress. Acta Hortic. 903:1187–1194.
- Lopez G, Behboudian MH, Girona J, Marsal J. 2012. Drought in deciduous fruit trees: implications for yield and fruit quality. In: Peter KV, editor. Plant responses to drought stress. Berlin/Heidelberg, Germany: Springer; p. 441–459. doi:10.1007/978-3-642-32653-0_17.
- Lourkisti R, Froelicher Y, Morillon R, Berti L, Santini J. 2022. Enhanced photosynthetic capacity, osmotic adjustment and antioxidant defenses contribute to improve tolerance to moderate water deficit and recovery of triploid citrus genotypes. Antioxidants. 562:11. doi:10.3390/antiox11030562.
- Mahouachi J, Gómez-Cadenas A, Primo-Millo E, Talon M. 2005. Antagonistic changes between abscisic acid and gibberellins in citrus fruits subjected to a series of different water conditions. J Plant Growth Regul. 24:179–187. doi:10.1007/s00344-004-0001-y.
- Maurin V, DesRochers A. 2013. Physiological and growth responses to pruning season and intensity of hybrid poplar. For Ecol Manag. 304:399–406. doi:10.1016/j.foreco.2013.05.039.
- Namirembe S, Brook RM, Ong CK. 2009. Manipulating phenology and water relations in senna spectabilis in a water limited environment in Kenya. Agrofor Syst. 75:197–210. doi:10.1007/s10457-008-9169-7.
- Pena-Rojas K, Aranda X, Fleck I. 2004. Stomatal limitation to CO2 assimilation and down-regulation of photosynthesis in Quercus ilex resprouts in response to slowly imposed drought. Tree Physiol. 24:813–822. doi:10.1093/treephys/24.7.813.
- Pope JO, Brown K, Fung F, Hanlon HM, Neal R, Palin EJ, Reid A. 2022. Investigation of future climate change over the British Isles using weather patterns. Clim Dyn. 58:2405–2419. doi:10.1007/s00382-021-06031-0.
- Rahmat BPN, Octavianis G, Budiarto R, Jadid N, Widiastuti A, Matra DD, Ezura H, Mubarok S. 2023. Sliaa9 mutation maintains photosynthetic capabilities under heat-stress conditions. Plants. 12:378. doi:10.3390/plants12020378.
- Roccuzzo G, Villalobos FJ, Testi L, Fereres E. 2014. Effects of water deficits on whole tree water use efficiency of orange. Agric Water Manag. 140:61–68. doi:10.1016/j.agwat.2014.03.019.
- Rodriguez J, Anoruo A, Jifon J, Simpson C. 2019. Physiological effects of exogenously applied reflectants and anti-transpirants on leaf temperature and fruit sunburn in citrus. Plants. 8:549. doi:10.3390/plants8120549.
- Romero-Trigueros C, Gambín JMB, Nortes Tortosa PA, Cabañero JJA, Nicolás EN. 2021. Isohydricity of two different citrus species under deficit irrigation and reclaimed water conditions. Plants. 10:2121. doi:10.3390/plants10102121.
- Sato K. 2015. Influence of drought and high temperature on citrus. In: Kanayama Y, Kochetov A, editor. Abiotic stress biology in horticultural plants. Tokyo, Japan: Springer. doi:10.1007/978-4-431-55251-2_6.
- Setiyoningrum F, Lioe HN, Apriyantono A, Abbas A. 2018. Drying and pulverization processes affect the physico-chemical properties of kaffir lime leaves (Citrus hystrix DC). Int Food Res J. 25:2620–2627.
- Shafqat W, Mazrou YSA, Rehman SU, Nehela Y, Ikram S, Bibi S, Naqvi SA, Hameed M, Jaskani MJ. 2021. Effect of three water regimes on the physiological and anatomical structure of stem and leaves of different citrus rootstocks with distinct degrees of tolerance to drought stress. Horticulturae. 7:554, 1–21. doi:10.3390/horticulturae7120554.
- Sovacool BK, Griffiths S, Kim J, Bazilian M. 2021. Climate change and industrial F-gases: a critical and systematic review of developments, sociotechnical systems and policy options for reducing synthetic greenhouse gas emissions. Renewable Sustainable Energy Rev. 141:110759. doi:10.1016/j.rser.2021.110759.
- Wongpornchai S. 2012. Kaffir lime leaf. In: Peter KV, editor. Handbook of herbs and spices. Cambridge, UK: Woodhead Publishing Limited; p. 319–328. doi:10.1533/9780857095688.319.
- Yuan R, Alferez F, Kostenyuk I, Singh S, Syvertsen JP, Burns JK. 2005. Partial defoliation can decrease average leaf size but has little effect on orange tree growth, fruit yield and juice quality. HortScience. 40:2011–2015. doi:10.21273/HORTSCI.40.7.2011.
- Zare H. 2021. Effects of different methods of pruning intensity on old fig (sabz cultivar) trees under rainfed conditions. Int J Fruit Sci. 21:379–391. doi:10.1080/15538362.2021.1892011.
- Zare H, Zare E, Sedaghat S, Jafari M. 2021. Investigation of different horticultural practices to minimize drought impacts in rainfed fig (Ficus carica L. ‘Sabz’). Acta Hortic. 1310. doi:10.17660/ActaHortic.2021.1310.32.
- Zhang X, Fan Y, Jia Y, Cui N, Zhao L, Hu X, Gong D. 2018. Effect of water deficit on photosynthetic characteristics, yield and water use efficiency in Shiranui citrus under drip irrigation. Trans Chinese Soc Agric Eng. 34:143–150. doi:10.11975/j.issn.1002-6819.2018.03.019.
- Zhou JJ, Zhang YH, Han Z, Liu XY, Jian YF, Hu CG, Dian YY. 2021. Evaluating the performance of hyperspectral leaf reflectance to detect water stress and estimation. Remote Sens (Basel). 13(11):2160. doi:10.3390/rs13112160.
- Ziogas V, Tanou G, Morianou G, Kourgialas N. 2021. Drought and salinity in citriculture: optimal practices to alleviate salinity and water stress. Agronomy. 11:1283. doi:10.3390/agronomy11071283.