
ABSTRACT
Spent mushroom substrate (SMS) is the residual biomass generated after harvesting the fruitbodies of edible/medicinal fungi. Disposal of SMS, the main by-product of the mushroom cultivation process, often leads to serious environmental problems and is financially demanding. Efficient recycling and valorization of SMS are crucial for the sustainable development of the mushroom industry in the frame of the circular economy principles. The physical properties and chemical composition of SMS are a solid fundament for developing several applications, and recent literature shows an increasing research interest in exploiting that inherent potential. This review provides a thorough outlook on SMS exploitation possibilities and discusses critically recent findings related to specific applications in plant and mushroom cultivation, animal husbandry, and recovery of enzymes and bioactive compounds.
HIGHLIGHTS
Valorization of spent substrate is crucial for a sustainable mushroom industry.
The review covers spent mushroom substrate (SMS) valorization for multiple uses.
SMS composition and mushroom species are essential factors for its exploitation.
SMS valorization forms an integral part of cascade use of plant biomass.
GRAPHICAL ABSTRACT
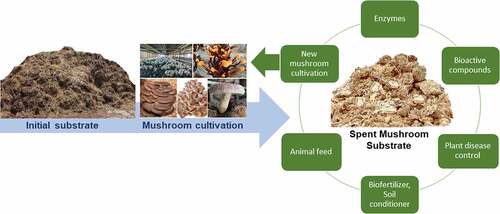
1. Introduction
Mushrooms are sporocarps, i.e. visible spore-bearing structures, fulfilling an essential function in the sexual reproductive stage of the life cycle of many fungi [Citation1]. Many mushrooms are considered edible because they do not contain toxins and are low in antinutrients, while they are rich in proteins, dietary fiber, vitamins, minerals, and other nutritional components [Citation2]. The specific composition of mushrooms depends on the species. Mushrooms can have up to 30% (w/w) crude protein, while the content of crude fiber, fat, and carbohydrates in some species can be up to 28, 8, and 95% (w/w), respectively [Citation3]. Edible mushrooms are climate-smart, protein-rich food sources that can partially substitute meat, whose production has a significant climate impact. Furthermore, due to their high content in various health-promoting ingredients, e.g. β-glucans, peptides, proteins, and phenolic compounds [Citation4], they possess immunomodulatory, antibacterial, cytostatic, antioxidant, and other properties, and for this reason, the term ‘medicinal mushrooms’ is also used when referring to them [Citation3].
The benefits of mushroom consumption on human health and wellbeing are well recognized. As a result, pertinent demand has considerably increased in all continents, and edible mushroom commercialization has nowadays become a worldwide business [Citation5]. Hence, mushroom production has increased more than 30 times since 1978, and it is a fast-expanding industrial activity. Although most of the production is concentrated in Asia, with China as the top producer with around 90% of the global market, mushroom production in the European Union, led by the Netherlands and Poland, and in the Americas has experiencing a significant increase during the last decades [Citation6]. The commercial cultivation of mushrooms includes more than fifty species. The top four belong to the genera Lentinula (L. edodes, popularly known as ‘shiitake’), Pleurotus (‘oyster mushrooms’), Auricularia (‘wood ear mushrooms’), and Agaricus (‘button mushrooms’), which together correspond to 74% of the world market [Citation7].
Mushrooms are cultivated on substrates based on plant biomass, e.g. crop residues and underutilized wood leftovers, which are continuously increasing because of the expansion of agricultural production driven by global population growth. Currently, disposal by burning is one of the chief methods for coping with the accumulation of plant residues. However, this widespread practice is against sustainability principles, contributes substantially to air pollution [Citation8], and results in a considerable waste of biomass resources that are highly valuable for generating materials, fuels, and chemicals of high economic and social value [Citation9].
The valorization of crop residues within new recycling models, i.e. substrates for mushroom cultivation, is crucial for the sustainability of agricultural production. Therefore, besides leading to the generation of food, mushroom cultivation is an example of holistic exploitation of residual lignocellulosic biomass through an efficient continuous-flow process carried out indoors, requiring remarkably lower land areas than most other crops [Citation10]. Furthermore, unlike conventional agriculture, which is season-dependent, mushroom production can be performed throughout the year independently of climatic conditions.
The mushroom cultivation process aims at producing fruitbodies of edible or/and medicinal fungi. At the end of the process, the fruitbodies are harvested, and an exhausted residual substrate is generated. That nutrient-depleted biomass waste, known as spent mushroom substrate (SMS), is the main by-product of the mushroom industry. Depending on the nature of the materials used for formulating the substrate, the type of production system, and the cultivated species, three to five kg of SMS is generated per kg of fresh mushrooms [Citation11]. In total, ca. 64 million tons of SMS were generated worldwide by the mushroom industry in 2018, and this figure could escalate to above 100 million tons by 2026 [Citation12].
The large quantities of generated SMS, currently regarded as a waste product with little inherent value, present a major challenge to mushroom producers due to the need to find suitable disposal sites and to cope with the high cost incurred for the transportation of a bulky material with high moisture content and low density; drying of fresh SMS is a hardly feasible energy-intensive activity. Moreover, SMS handling/disposal is of primary environmental concern due to the emission of greenhouse gases from spontaneous anaerobic digestion (often occurring in the piles formed during provisional storage), foul odors, and leachate drainage to water receptors causing pollution and eutrophication [Citation13]. Landfilling has traditionally been the chief disposal strategy for SMS, but it is now banned in the European Union by a Council Directive on landfilling of biodegradable wastes [Citation14]. The current linear ‘take, make, dispose of’ approach, where SMS is regarded as waste, threatens the future development of the mushroom-growing sector. Valorization of SMS is crucial for developing a sustainable mushroom industry in the frame of a circular-economy model. It is essential to investigate SMS characteristics to identify appropriate valorization alternatives.
SMS composition and properties are mainly associated with the type of raw materials and supplements used to prepare the initial mushroom substrate. For the cultivation of edible mushrooms of the genera Lentinula, Pleurotus, and Auricularia, which represent 60% of the global production, various lignocellulosic by-products, e.g. forest, agricultural and agro-industrial residues, are used as substrate base. Chicken manure is also a major component for other mushroom species requiring composted substrates (e.g. those of the genus Agaricus). Starch-containing and nitrogen-rich ingredients (e.g. cereal bran or legumes’ flour) and mineral salts are used as supplements. During cultivation, substrate components are enzymatically degraded, and the resulting nutrients (together with others existing in the substrate) are used for fungal growth and mushroom production. Mass losses in the ranges of 26–46%, 57–77%, and 61–75% of the initial cellulose, hemicelluloses, and lignin, respectively, have been reported for Pleurotus ostreatus, Pleurotus pulmonarius, and L. edodes [Citation15–17]. In the end, SMS composition strongly depends on the nature of the initial substrate and the cultivated species [Citation18]. Therefore, SMS primarily consists of plant cell-wall components (lignin, hemicelluloses, cellulose) and residual fungal mycelium, as well as non-cell-wall carbohydrates, proteins, and minerals.
There are different valorization routes for SMS, and some of them have already been discussed in previously published reviews [Citation19,Citation20]. The current review is aimed at providing, in brief, an updated overview of potential SMS applications and products related to (i) new cycles of mushroom cultivation, (ii) agriculture and animal husbandry, and (iii) the production of enzymes and bioactive compounds. SMS valorization as part of cascade-use systems for plant biomass processing is also discussed. Bioremediation and energy-related uses are not included because they were exhaustively presented in a recent review [Citation20]. This review is based on an exhaustive Scopus search performed in July 2022. The search terms used were Spent Mushroom Substrate OR Spent Mushroom Compost AND relevant keywords of each specific application. The topic presented in this review is of relevance to the UN Sustainable Development Goals 2 (Zero hunger), 3 (Good health and well-being), 9 (industry, innovation, and infrastructure), 13 (climate action), and 15 (life on land), considering that the discussed valorization alternatives have the potential for providing innovative solutions to increase food security, and contributing to the production of healthy food and reduction of the use of harmful chemicals in farmlands.
2. Reusing spent mushroom substrate for new cultivation of mushrooms
The spent mushroom substrate can be used in substrate formulation for new cycles of mushroom cultivation provided that suitable lignocellulosic materials are employed, the fungal strain is appropriately selected, and the environmental conditions are optimally regulated. Supplemented cereal straw and wood sawdust are the most common substrates in commercial mushroom cultivation due to their composition, availability, and relatively low cost. Agricultural or agro-industrial by-products with low or no economic value, such as sugarcane bagasse, coffee husks, and olive mill and winery wastes, are exploited in mushroom production, contributing to both the improvement of cultivation performance and the enhancement of mushrooms nutritional value [Citation21–24]. Using cheap lignocellulosic residues positively affects the cost of substrate, providing an environmentally friendly solution for their effective management and valorization.
Cultivated mushrooms are often grouped – based on their ecological adaptation/requirements – as either primary decomposers (e.g. P. ostreatus, L. edodes) that are produced directly on previously untreated (or partly treated/composted) lignocellulosic substrates, or as secondary decomposers (e.g. Agaricus bisporus, Volvariella volvacea). Secondary decomposers are cultivated on composted substrates prepared from various agricultural residues, including manures. The proposal to reuse SMS in new crops was originally based on the sequential use of the substrate, first by primary decomposers and then by secondary decomposers, and on the enzymes involved in each process since these vary among species of different ecological groups [Citation20,Citation25]. However, in most studies, supplementation is required to adjust the nutrient content when SMS is used as the sole (or the main) substrate ingredient in mushroom cultivation. Hence, this material could be exploited to cultivate a broader range of mushroom species (not only secondary decomposers). Furthermore, many of the most successful SMS applications have been reported when the same species (as the one originally cultivated on the spent substrate) was also used in the new crop, e.g. P. ostreatus, Auricularia polytricha, A. bisporus [Citation26–29]. The initial substrate composition, the cultivation cycle duration, and the number of flushes harvested are important in optimizing SMS for reuse in mushroom cultivation. The type of substrate pretreatment adopted (e.g. chopping, composting) prior to cultivation, the incorporation rate of SMS to the main substrate of the new crop, further supplementation with nutrients, and the selection of the species/strain to be used are also important parameters, which have to be considered when such types of applications are developed. Factors affecting the success of new mushroom crops based on SMS recycling are summarized in .
Figure 1. Factors affecting cultivation parameters and the use of SMS in new mushroom crops.

Reported results from using SMS in new mushroom crops demonstrate a wide variation as regards the effect of the recycled material on the final yield (). In several cases, similar [Citation27,Citation29,Citation40,Citation44,Citation50] or even higher [Citation26,Citation28,Citation45] values of biological efficiencies (ΒΕ; percentage ratio of fresh mushroom weight over the dry weight of the respective substrate) were recorded in substrates containing SMS deriving from the cultivation of either the same or other mushroom species when compared to conventional (used for the first time) substrates. However, in some studies, it was also reported that the incorporation of high amounts of SMS in the new cultivation medium or the casing layer negatively affected the mushrooms’ final yield [Citation30,Citation36,Citation37,Citation42], which could be mainly attributed to the low content of nutrients or to inadequate supplementation of the spent substrates.
Table 1. Reuse of spent mushroom substrate (SMS) for the cultivation of various mushroom species as reported in pertinent publications: origin and composition of SMS, mushroom to be cultivated, new substrate formulation and supplements, biological efficiency (BE) reported for the crop obtained and main comments on the results of the respective study. Abbreviation used: NR, not reported.
Most of the investigations related to the reuse of SMS in new cultivation cycles focus on species of the genus Pleurotus. Indicatively, out of the 27 selected studies shown in , 16 deal with the reuse of Pleurotus SMS, and 14 with the cultivation of oyster mushrooms in SMS-containing substrates. This may be explained by the relative ease of oyster mushroom cultivation, their rather short production cycle, and the wide range of suitable substrates available. Among the most relevant examples are those pertaining to the use of Pleurotus eous SMS mixed with wheat straw for the cultivation of other Pleurotus species (BE up to 113% [Citation45]) and the use of supplemented SMS from Pleurotus sajor-caju for the production of the same mushroom (BE up to 125% [Citation47]). Another successful example is the use of supplemented SMS from P. ostreatus as substrate for growing P. ostreatus and P. pulmonarius mushrooms, which resulted in the highest BE values reported in pertinent literature, namely 185% for P. ostreatus and 208% for P. pulmonarius [Citation26]. Furthermore, Pleurotus SMS can also be exploited in crops of other species providing satisfactory yields, as it is the case in Lyophyllum decastes, where the use of P. ostreatus SMS with bark compost and rice straw provided BE of 134% [Citation51]. In contrast, cultivation of Pleurotus species on spent substrates from other mushrooms, i.e. Pholiota nameko, Hypsizigus marmoreus, and L. edodes exhibited substantially lower BE (23–62%) [Citation36,Citation46,Citation50].
Shiitake (L. edodes), a widely cultivated edible mushroom, is produced mainly on hardwood sawdust substrates. SMS deriving from L. edodes seems to be suitable for the cultivation of various oyster mushroom species, including P. ostreatus, P. sajor-caju and Pleurotus cornucopiae (BE: 61–79%) as well as for Flammulina velutipes (BE 88%) following rich supplementation with cereal derivatives [Citation50,Citation52]. On the other hand, using SMS from other mushrooms (e.g. A. bisporus and F. velutipes) to cultivate shiitake requires mixing with untreated sawdust at a rate of at least 40% [Citation48,Citation49].
The reuse of SMS deriving from the cultivation of Agaricus species to establish new mushroom crops is quite demanding due to the nature of the final material and the processes leading to its production. Therefore, most attempts focus either on suitably upgrading it with soybean meal and Target® (a commercial delayed release nutrient suitable for mushroom cultivation) before reusing it as an ingredient for A. bisporus cultivation (ΒΕ: 97–144%) [Citation27]. Attempts focusing on exploiting SMS as casing material alone or mixed with farm yard manure or sphagnum peat in the cultivation of Pleurotus eryngii and A. bisporus, respectively, have also been reported [Citation38,Citation42].
Concerning the use of SMS of less widespread species, noteworthy is the case of F. velutipes SMS – which when combined with oak sawdust and rice bran – exhibited satisfactory yields of L. edodes mushrooms (BE: 60–84%) [Citation48]. In addition, the sawdust-based SMS from Ph. nameko, H. marmoreus and Hericium erinaceus were used to produce P. ostreatus mushrooms (BE: 66–73%) [Citation30].
The reuse of SMS in new mushroom crops seems to have considerable potential since it can support high yields and is both financially feasible and environmentally sustainable. The elements and organic compounds existing in SMS constitute valuable sources of energy and nutrients, which can partially or entirely cover the needs of additional cultivation cycle(s) after suitable treatment or supplementation.
3. Spent mushroom substrate as feed
It is estimated that agricultural production should be increased by 70–100% to meet the food demand of the increasing global population, which is predicted to grow to 9.7 billion by 2050 [Citation53]. Soybeans and maize are the most common energy and protein sources used by livestock farmers to generate meat, eggs, and dairy products, which, in turn, are the main protein sources in human diet [Citation54]. The need for animal feed production is predicted to increase significantly, and the feed industry must look for additional/alternative means to cover the respective demand. Exploiting suitable bioresources (e.g. SMS) could contribute toward this direction by readily providing material to be used as feed supplement.
The main raw materials used in mushroom cultivation are rich in cellulose, hemicelluloses and lignin, while their protein content is generally low [Citation23]. During solid-state fermentation by mushroom-forming fungi, the substrate polymers are enzymatically degraded, and the digestibility of plant residues is considerably improved. Concomitantly, the growth of mycelial biomass upgrades the substrate by increasing its content in proteins and bioactive compounds, e.g. polysaccharides and ergosterol [Citation55–58]. Indicatively, the growth of P. ostreatus mushrooms on faba bean hulls increased their protein content from 208 g kg−1 (on a dry weight (DW) basis) in the initial substrate to 347 g kg−1 in the SMS [Citation59]. Furthermore, P. ostreatus growth enriched the material in 14 out of 16 analyzed amino acids, and significantly reduced the content of anti-nutritional compounds, such as tannin, vicine, and convicine. SMS from other mushroom species is also rich in compounds of interest for enhancing the quality of feed rations. L. edodes SMS is rich in the provitamin D2 ergosterol (151.6 mg ergosterol equivalents/100 g) [Citation58], while SMS from several other species contains high amounts of polysaccharides, including β-glucans [Citation60]. Consequently, the high nutritional value of SMS is the main factor for its inclusion in the diets of poultry, ruminants, and monogastric animals, and, recently, in fish and edible insects. A summary of relevant reports on the reuse of SMS as animal feed is presented in . In addition, the main outcome of each study is briefly presented and further discussed below.
Table 2. Reuse of spent mushroom substrate (SMS) as animal feed based on the outcome reported in pertinent publications. Abbreviations used: ADF, acid detergent fiber; ADL, acid detergent lignin; HWE, hot water extract; NDF, neutral detergent fiber; NR, not reported; S-SMS: steam-treated SMS; UN-SMS: untreated SMS.
3.1. Spent mushroom substrate in the diet of poultry
The incorporation of SMS derived from the cultivation of P. eryngii, P. ostreatus and H. marmoreus (fermented or not by Bacillus subtilis) at a ratio from 5 to 15% (w/w) in a poultry diet increased the feed intake without having adverse effects on the egg production and the mass of useful meat [Citation67,Citation74,Citation75,Citation79]. On the other hand, incorporation of Agaricus blazei SMS at rates exceeding 0.4% (w/w) caused a gradual reduction in the weight gain of broiler chickens, while inclusion ratios of only 0.2% exhibited the highest value of weight gain and feed intake, as well as the best feed conversion [Citation81]. Similarly, low inclusion ratios of P. sajor-caju SMS (up to 0.67%) improved the weight gain of broiler chicks in the first 21 days [Citation80].
3.2. Spent mushroom substrate in the diet of monogastric animals
SMS inclusion in the diet of monogastric animals have been tested with both pigs and model animals (mice, rats). The addition of sawdust-based SMS from Grifola frondosa (25% w/w) in rats’ diet did not affect the weight gain, the feed efficiency, or the biochemical parameters, while fecal weight and protein content were found to be higher [Citation76]. In addition, the – orally administered – hot water extract of SMS from Ganoderma lucidum exhibited enhanced murine immune function in mice [Citation65]. Furthermore, the use of low amounts of SMS from P. ostreatus (up to 3.5%, w/w), Cordyceps militaris (0.2%, w/w), and L. edodes (3%, w/w) in pigs’ diet positively affected the feed intake and conversion, as well as the final weight and quality of meat deriving from trials [Citation55,Citation64,Citation82]. C. militaris SMS resulted in increased immunoglobulin A and G, and glutathione peroxidase activities, while leukocytes, cholesterol and malondialdehyde contents were decreased [Citation55]. Similarly, beneficial effects on the intestinal mucosal barrier, immunity, and the diversity and abundance of the bacteria in the colon and cecum were observed for weaned piglets when fed with L. edodes SMS [Citation64]. It is noteworthy that SMC seems to be useful as a ‘behavior regulator’ in pigs; by having access to mushroom compost through a metal grid, pigs demonstrated significantly reduced negative behavior (e.g. such as nosing, tail biting and chewing) against penmates, as well as improved overall welfare in comparison to pigs with no access to SMC [Citation84].
3.3. Spent mushroom substrate in the diet of ruminants
The use of SMS as animal feed has been investigated to a larger extent for ruminants than for monogastric animals (i.e. 13 vs. 5 publications appeared, respectively, when a Scopus search was performed by using the keywords ‘Spent Mushroom substrate/compost’ AND ‘feed;’ July 2022). Incorporating SMS from various mushroom species at a rate of up to 30% (w/w) in the daily intake of ruminants revealed its potential as a supplement to conventional feeds without affecting several relevant parameters (). Specifically, feeding sheep for three weeks with a diet including up to 20% (w/w) of A. bisporus SMS did not affect the nutrient intake, digestibility, and nitrogen balance [Citation83]. Similarly, A. bisporus SMS fed at a rate of 15% (w/w) for 170 days did not cause any effect on the carcass and internal organs of Holsteins male calves [Citation73]. Plain P. ostreatus SMS in ratios higher than 15% resulted in adverse effects in sheep slaughter weight, empty body weight, and hot and cold carcass weight [Citation69]. In contrast, when rice straw was fermented for eight weeks with P. sajor-caju SMS before being fed to alpine dairy goats, it increased the rumen degradable fibers fraction and improved dry matter intake and milk yield [Citation61].
Feeding male sika deer for 60 days with P. ostreatus SMS (10%, w/w) resulted in a reduction of the intake of organic matter and improved digestibility of crude fibers, while no effect on either the apparent nutrient digestibility, feed intake, velvet antler production, or biochemical indexes was observed when F. velutipes SMS (10%, w/w) was fed to the same animals [Citation62]. When hot water extracts from G. lucidum and Ganoderma chalceum (syn. G. balabacense) SMS were supplemented to dairy cow feed, immunity and antioxidant capacity were increased, and milk quality was improved [Citation71,Citation72]. By using SMS extracts, the addition of large amounts of fibrous components from the untreated SMS could be avoided, but further studies are required to investigate their impact on animal health and the optimum incorporation rate, which depends mainly on the substrate origin.
Agaricus and Pleurotus species are usually cultivated in straw-based substrates, while L. edodes, G. lucidum, Gr. frondosa and He. erinaceus are cultivated in wood-based substrates. The incorporation rate of such substrates in animal feed is low, and further treatment is necessary to improve their nutritional characteristics. In recent years, microbial fermentation with probiotic microorganisms has been adopted as a cheap, fast, and efficient method to reduce fibrous ingredients and upgrade the protein content of SMS, including those deriving from sawdust-based media. Moreover, probiotic microorganisms relieve animals weaning stress, regulate intestinal microbiota, and reduce diarrhea incidents [Citation85,Citation86].
Due to its high moisture content, SMS tends to decompose rapidly; hence, it needs to be processed quickly. This could be achieved by ensiling, for instance, by lactic acid fermentation under anaerobic conditions [Citation87]. Lactic acid bacteria produce desirable metabolites, and suppress the growth of clostridia and other deleterious microbial populations [Citation88]. Although ensiling processes may be initiated naturally by the epiphytic microorganisms existing in the initial material, they can be assisted by inoculated bacteria. Inoculation of SMS with Lactobacillus, Bacillus, or Enterobacter spp. ensures rapid acidification, and increases dry matter degradability and crude protein content [Citation64,Citation66,Citation70,Citation75,Citation77,Citation78].
An indicative example is the use of a sawdust-based P. eryngii SMS incorporated at a high rate (45%, w/w) into silage with various agricultural by-products, and fermented for 22 days [Citation68]. Using the resulting product for feeding sheep resulted in similar energy value, lower digestion of fibers, and higher protein metabolism and utilization compared to the outcome achieved with a rye straw-based control diet. In addition, the sawdust-based SMS from the same mushroom species, when fermented with Enterobacter and Bacillus spp., significantly improved growth performance and carcass traits in Hanwoo steers compared to rice straw feed administered for 12.6 months during the growing and fattening periods [Citation77]. Similarly, P. ostreatus SMS fermented with Lactobacillus plantarum and Pediococcus acidilactici could replace up to 50% of the conventional feed provided to Hanwoo steers and postweaning calves [Citation66,Citation70,Citation78]. Such an SMS-based feed improved the growth performance of the tested animals or enhanced the daily gain caused by increased voluntary feed intake. Finally, feeding Liuyang black goats with P. ostreatus SMS co-fermented with whole rice plants improved meat quality and had no adverse effects on the slaughter performance [Citation63].
3.4. Spent mushroom substrate in the diet of fish and edible insects
Using SMS in pisciculture is also of substantial interest. SMS from P. ostreatus, Pleurotus cystidiosus, and G. lucidum seems to support the growth of catfish, and significantly increase its survival rate and digesting ability compared to commercial feeds [Citation89]. The addition of C. militaris SMS up to 40 g kg−1 in the diet of Nile tilapia (Oreochromis niloticus) improved growth performance, skin mucus lysozyme, and peroxidase activities, as well as serum immune parameters [Citation90]. The combination of SMS with L. plantarum further improved those parameters. Moreover, enrichment of Nile tilapia diet with A. blazei SMC (1%, w/w) provided significant protection against infections from Streptococcus agalactiae [Citation91]. Including an extract from the SMS of Schizophyllum commune, a popular mushroom in Thailand, in the feed of Nile tilapia resulted at enhancing their immune defense [Citation92].
In the frame of the need to reduce dependence on feeds deriving from plants, the use of insects seems to be a promising alternative due to their high content in crude protein (up to 76%) and fat (up to 59%), energy (20–30 MJ/kg DM), as well as to their short life-cycle and the low-cost growth prerequisites [Citation93–95]. The potential of six SMS derived from Auricularia cornea, Auricularia heimuer, P. eryngii, P. ostreatus, Pleurotus citrinopileatus, and L. edodes was recently evaluated to rear black soldier fly (Hermetia illucens) and Tenebrio molitor larvae. L. edodes SMS was shown to be the most suitable to replace the insects' conventional feed [Citation96,Citation97]. Furthermore, when Protaetia brevitarsis larvae were grown on L. edodes or Auricularia auricula-judae SMS, a nutrient-rich organic fertilizer with low phytotoxicity and high humic acid content was produced [Citation98]. However, no studies exist on the production of insects naturally feeding on mushroom substrates. That could be a promising alternative considering the ease of insects’ growth and the low demand in terms of material resources.
In conclusion, using SMS in animal nutrition can significantly contribute to the enrichment of feed, particularly regarding proteins and bioactive compounds. However, incorporating SMS into the daily feed-schedule is a complex process. The mushroom species, the initial substrate composition, the animal species, and the digestibility and voluntary intake of the final product are factors that must be carefully considered to calculate the final integration rate. The high NDF and ADF content (especially in sawdust-based substrates) is probably the main limiting factor in SMS exploitation as feed. Adopting appropriate treatment approaches, including lactic acid fermentation and the use of SMS extracts, could enhance the nutritional and acceptance characteristics, thus facilitating the incorporation of SMS in the diet of productive animals. Particularities related to the composition of each type of SMS and the individual needs of the animal species require careful experimentation on a case-by-case basis to ascertain the safe and efficient use of SMS.
4. Use of spent mushroom substrate in agriculture
The global demand for food and feed has led to intensification of agricultural production and the widespread use of fertilizers and pesticides. World consumption of the three main fertilizer elements (Ν, P, K) was estimated at 201.7 million tons in 2020 [Citation99], and nearly 3 billion kg of pesticides are used yearly [Citation100]. Although using fertilizers and pesticides has increased food availability, their extensive application negatively impacts the environment and human health. Hence, adopting sustainable agronomic practices, including the development of novel environment-friendly and cost-effective biofertilizers and biopesticides, is of high priority. In line with that approach, the SMS’s physical properties, its high content of bioactive compounds, and readily available macro- and trace elements make it a promising candidate for several agricultural applications, the most important of which are presented in the following paragraphs.
4.1. Use of spent mushroom substrate as biofertilizer and soil conditioner
Organic soil amendments, commonly used in agriculture, exert positive effects on crop productivity and soil health by affecting physicochemical and biological properties of soil [Citation101–103]. Among the most widespread materials used as organic soil amendments are those originating from municipal wastes (food and gardening wastes, sewage sludge), animal husbandry (manure), crop production (stems, leaves and branches), and agro-industrial activities (fruit pulp and oil extraction by-products). However, those materials can contain hazardous compounds or plant pathogens, which are detrimental to soils and crops.
Since SMS is rich in nutrients and has low (and, most often, no) content in xenobiotic compounds and heavy metals, it can be used as a soil amendment either directly or after a composting treatment. SMS properties vary depending on the raw materials included in the initial substrate, the mushroom species, and the cultivation technology. Accordingly, a wide range of effects is noted on soil characteristics, crop growth and yield when SMS is used as a soil conditioner or fertilizer [Citation104,Citation105]. However, it is worth mentioning that the mushroom species and the SMS composition are often not specified in pertinent publications, making it difficult to draw sound conclusions about its exploitation prospects.
A summary of relevant reports on incorporating SMS into soils is presented in , which includes information on the SMS origin, type, and incorporation rate, and the main effects of SMS addition on the soil and plants under study. The presented results indicated improvements in soil structure and fertility, which led to increased crop production or contributed to the restoration of barren lands and degraded soils.
Table 3. Reuse of spent mushroom substrate (SMS) as soil amendment based on the outcome reported in pertinent publications. Abbreviations used: NPK, nitrogen, phosphorus, potassium NR, not reported; OC, organic carbon; OM, organic matter; PGPB: plant growth promoting bacteria; TN, total nitrogen.
By applying SMS of unknown origin (20 Mg ha−1) and chicken manure (10 Mg ha−1) in a sandy soil every one-two years for 20 years, Lipiec et al. [Citation109] reported an increase in soil organic matter content by 102–201%. The experiment also resulted in a long-term increase in field water capacity caused by the augmentation of residual pores by up to 251%. Similarly, fresh or composted SMS applied annually for four years at two different rates (8 and 25 Mg ha−1) to a semiarid vineyard soil increased the content of inorganic N in the soil surface (0–5 cm) [Citation120]. However, only the highest SMS addition rate improved soil organic carbon, total nitrogen, and labile organic forms at 0–5 and 5–15 cm soil depths.
In other large-scale applications, incorporating A. bisporus SMS into the soil (100 kg ha−1) increased oxidizable organic carbon, organic N, and available P content [Citation119]. The values obtained for using A. bisporus SMS alone were higher than those resulting from incorporating a mixture of A. bisporus and P. ostreatus SMS (1:1, v/v). Both schemes of SMS addition resulted in increased phosphatase activity compared to unamended soil, while no alterations in the soil salinity or pH value were observed, and N mineralization was low. The same treatments also had positive effects when examined in a calcareous clayey-loam soil used for lettuce production [Citation105]. In that study, application of SMS resulted in higher values of oxidizable organic carbon, organic N, extractable K, available P, and cation exchange capacity (especially when using A. bisporus SMS) than in soils receiving NPK fertilization, while lettuce yields were similar.
Ngan and Riddech (2021) reported the application of a mixture of SMS with plant growth-promoting bacteria (Bacillus amyloliquefaciens) in the cultivation of Hibiscus sabdariffa [Citation110]. The study revealed an improvement in soil properties exceeding the effect exerted by NPK fertilization. Unfortunately, the lack of information on the SMS origin makes it difficult to compare the results with those of other relevant studies.
Testing fresh or sterilized F. velutipes SMS in cucumber cultivation resulted in a significant increase in total organic carbon, dissolved organic carbon, and microbial biomass carbon compared to NPK use and to no fertilization [Citation111]. The study revealed higher levels of microbial diversity and enzyme activities for the fresh SMS-amended soil compared to soil treated with mineral fertilizer. Correspondingly, A. bisporus SMS amendment in soils increased bacteria and fungi co-occurrence, and the plant yield was positively affected by the relative abundance of microbial hubs [Citation112]. Similarly, the application of Agaricus subrufescens and L. edodes SMS enhanced soil microbial population, and resulted in a remarkable increase in lettuce plants' dry weight compared with the results achieved with no fertilization or NPK treatments [Citation104]. For several other crops, SMS application to the soil led to higher yields than those obtained by mineral fertilization [Citation105,Citation116,Citation121].
Soil biological properties play a critical role in the maintenance of ecosystem functions, crops productivity enhancement, and at mitigating the adverse effects of pollutants. The beneficial effect on soil biological properties, including the structure of microbial communities and associated enzyme activities, is an attractive aspect of using SMS as an amendment. The main disadvantage of SMS is the state of stability/maturity which – if imperfect/immature – could hamper its wide agronomic use. However, this issue could be overcome by composting it, alone or mixed with other crop residues, under controlled conditions [Citation115].
Several studies have revealed that using SMS as an ingredient in the composting process promotes the degradation of organic matter in mixtures with waste sludge, pig manure, corn stalks, and cow dung [Citation122–124]. It has also resulted in the enhancement of the humification process [Citation125], at reducing ammonia emissions [Citation122,Citation126], facilitating heavy metal passivation [Citation125], and improving the quality of the final product [Citation122,Citation124]. Furthermore, using composted A. bisporus SMS alone as a substrate for the cultivation of Lolium multiflorum resulted in a yield improvement by up to 300% compared to the reference of NPK fertilization [Citation115]. Co-composting of Au. auricula-judae SMS with biogas residues and pig manure led to the production of higher quality seedlings than those obtained from commercial substrates [Citation113]. Substrates containing composted A. bisporus and P. ostreatus SMS resulted in increased yields of baby leaf lettuce, even in the presence of the soil-borne plant pathogen Pythium irregulare [Citation108]. Adopting appropriate methodologies, such as the addition of enzymes or earthworms (vermicomposting), during the composting process should result in further improvement of the quality of composted SMS by promoting the beneficial effects of autochthonous bacteria, increasing ion-exchange capacity, decreasing total carbon and C/N ratio, and promoting the synthesis of nitrates [Citation127–129].
In conclusion, the use of SMS as soil amendment has beneficial effects on soil fertility and structure. SMS presents a promising potential for substituting, at least partially, the use of mineral fertilizers in continuous crops and thus contributes at mitigating soil secondary salinization and acidification, and at avoiding nutrient imbalances and accumulation of toxic allelochemicals.
4.2. Use of spent mushroom substrate for plant-disease control
To deal with the negative repercussions of using chemical-based pesticides in agriculture, the application of environmentally friendly products for pest protection is crucial. Biocontrol agents, including live organisms and biological pesticides, are potential alternatives for controlling plant diseases. In contrast to chemical pesticides, biocontrol agents have little impact on non-targeted organisms; they do not leave behind any long-lasting harmful leachates and do not lead to the development of resistant microbial strains or insects. However, they often exhibit low-medium effectiveness and a shorter shelf life [Citation130,Citation131].
The bioactive compounds in SMS have antimicrobial properties [Citation132], which could be exploited against plant pathogens. Although in vitro studies have shown the potential suitability of mushroom and mycelium extracts against plant pathogens [Citation87,Citation133,Citation134], they do not necessarily reveal the in vivo effectiveness. SMS application has shown to be effective in suppressing plant disease incidence. shows examples of reported research findings on using SMS for controlling plant pathogens and pests, and also includes the SMS origin, the plant – pathogen/pest system, and the main outcome of each study.
Table 4. Reuse of spent mushroom substrate for the control of plant pathogens and pests based on the outcome reported in pertinent publications. Abbreviations used: ACT, aerated compost tea; CT, compost tea; NCT, non-aerated compost tea; NR, not reported.
Several studies on SMS-based biocontrol products against plant diseases concern L. edodes. The in vitro antimicrobial activity of L. edodes SMS [Citation87,Citation151], was further evidenced when hot water extracts were used to inhibit the germination of Pyricularia oryzae conidia in rice plants and to suppress the growth of Phytophthora capsici in pepper plants [Citation141,Citation142]. A chitin/cellulose nanofiber complex isolated from L. edodes SMS exhibited significant activity against Alternaria brassicicola in Arabidopsis thaliana plants [Citation135]. L. edodes SMS-based biocontrol agents reduced the disease symptoms and promoted plant growth [Citation135,Citation142].
P. ostreatus SMS can provide another alternative to suppress plant diseases. Paddy straw-based P. ostreatus SMS, bio-fortified with Trichoderma asperellum, led to a remarkable reduction of the severity index of Fusarium oxysporum-induced disease, while it contributed to enhanced tomato growth [Citation137]. The application of a polysaccharide extract from P. ostreatus SMS and discarded L. edodes mushrooms reduced by 50% the severity of bacterial spot caused by Xanthomonas gardneri in tomato cotyledons, leaflets, and five-leaf plants [Citation139]. Phenolic-rich extracts from P. ostreatus SMS have been shown to prevent the development of the parasitic plant broomrape in faba bean cultivars [Citation152], and to improve the rice growth and yield parameters [Citation153]. In another study, mixing composted SMS from either P. ostreatus or V. volvacea with a biofertilizer exhibited higher control efficacy against Ralstonia wilt and Phytophthora blight diseases, than using the biofertilizer alone [Citation138].
SMS from less widely cultivated mushrooms has also shown suppressive activity against plant diseases. Application of SMS from Ly. decastes and P. eryngii into soils used for cultivating cucumber resulted in protection against disease symptoms caused by Colletotrichum orbiculare, Podosphaera xanthii, Cladosporium cucumerinum and Pseudomonas syringae [Citation147,Citation149]. A protective effect against Colletotrichum lagenarium in cucumber plants was observed after spraying a water extract of Ly. decastes SMS [Citation149]. The incorporation of Ly. decastes SMS into soil suppressed the lesions caused by Al. brassicicola in Arabidopsis thaliana leaves; this effect was attributed to SMS volatile components [Citation136].
The results so far indicate that SMS richness in antimicrobial compounds in concomitance with its natural microbiome, including organisms suppressing soil-borne plant pathogens, are essential prerequisites for developing relevant plant-disease control products. However, further experimentation, including evaluation in large-scale greenhouse and field trials, is required to fully benefit from that potential toward a solid sustainable agriculture model.
4.3. Effects of SMS on nutritional value and secondary metabolites production in plants
Plant secondary metabolites, including vitamins, terpenoids and polyphenols, in fruits and vegetables are important for reducing risks of cardiovascular diseases and maintaining good health [Citation154,Citation155]. Those molecules exert a wide range of effects on the plant and associated organisms, and their production depends on various biotic and abiotic factors [Citation156].
SMS application affects the content of secondary metabolites in plants. Vahid Afagh et al. [Citation157] reported that the incorporation of Agaricus SMS leachates in sandy soil (up to 15% (v/v)) significantly increased the content of essential oil, proline, and soluble sugars in chamomile (Matricaria chamomilla) in comparison to plants grown on non-supplemented soil. Increasing the SMS leachate content enhanced K and Na absorption, whereas N and P uptake was not affected. Similarly, the addition of SMS leachate (20–60% (v/v)) in the soil led to increased content of essential oil components, chlorophyll, and antioxidant compounds in chamomile [Citation158]. Application of SMS as an amendment in soils where basil (Ocimum basilicum) was cultivated, resulted in a two-fold increase in essential oil components, and in an enhancement of its content in micro- and macronutrients [Citation159].
SMS use in the cultivation of vegetables demonstrated a wide range of effects on the various parameters, including product yield and quality. Applying a leachate of P. ostreatus SMS and A. bisporus SMS (10–25% (w/w)) to the soil increased the content of chlorophyll in pepper leaves, and that of carotenoids and protein in fruits [Citation160]. Furthermore, A. bisporus SMS biofortified with Trichoderma harzianum inhibited lipid peroxidation and protein oxidation with a significant increase in total polyphenol and flavonoid contents in tomatoes, and enhanced Fe2+/Fe3+ chelating activity and superoxide anion radical scavenging activity compared to an SMS-free control [Citation161]. Similarly, P. ostreatus SMS biofortified with Trichoderma asperellum improved morpho-biochemical and nutritional parameters, such as the content of chlorophyll, carotenoids, total soluble sugars, total soluble proteins, lycopene, β-carotene, and ascorbic acid, and antioxidant properties, of tomato plants [Citation137]. Another study, using SMS from A. bisporus or P. ostreatus for replacing peat moss by 25–100% (w/w), reported that the effect of SMS on the macronutrient content of tomato, courgette, and pepper plants was species-dependent [Citation162]. A proportional increase of N content with the increase of SMS ratio in the substrate was observed for pepper, whereas no significant effect was evident for courgette and tomato. In addition, increasing the incorporation volumes of SMS increased K content for courgette and pepper, but not for tomato. Last, courgette and pepper exhibited similar P content when grown on SMS-based substrates and a peat control, whereas P content in tomato seedlings grown on SMS-based substrates was lower than in plants grown on peat.
Although the scientific data on the effects of SMS on the nutritional value of edible and medicinal plants are still limited, the available results reveal SMS potential to increase the content of specific elements and secondary metabolites in plants.
5. Spent mushroom substrate as source of enzymes and bioactive compounds
Producing enzymes and different bioactive compounds is a reasonable way of SMS valorization. SMS-derived enzymes are of interest in industrial sectors, such as brewing, baking, starch-processing, leather, and textile industries, as well as in bioremediation and the emerging biofuel and biorefinery business. SMS-derived bioactive molecules have also the potential for application in the pharmaceutical, biomedical, feed, and food sectors.
5.1. Enzymes
SMS is a source of various enzymes that can be recovered by extraction with different solvent systems. Furthermore, SMS can be used as substrate for the cultivation of enzyme-producing microorganisms.
5.1.1. Recovery of enzymes from spent mushroom substrate
For growing on lignocellulosic biomass, white-rot fungi secrete hydrolytic and oxidative enzymes responsible for degrading complex polymers into low-molecular weight substances, which can be assimilated for fungal growth [Citation163]. The main groups of enzymes participating in fungal degradation of lignocellulosic materials are presented in . Hydrolytic enzymes are responsible for deconstructing cellulose and hemicelluloses, while oxidative enzymes are involved in lignin degradation [Citation7]. Consequently, upon the end of cultivation, SMS contains extracellular fungal enzymes, such as ligninases, cellulases, and hemicellulases, that can be recovered using different extraction procedures. The level of enzyme activities and their corresponding titers depend on the growth substrate and the fungal species’ ability to degrade different lignocellulose components. For example, since white-rot fungi degrade lignin and hemicelluloses preferentially, extracts of their spent substrates are rich in ligninases and xylanases, while cellulase activity is hardly detected.
Figure 2. Enzymes participating in fungal degradation of lignocellulosic substrates.

The enzymatic systems present in SMS of various fungal species make possible their application for different purposes. For example, P. ostreatus SMS can be applied for decolorizing textile effluents because it contains oxidoreductases that degrade the dye molecules [Citation164]. Similarly, the laccase and manganese peroxidase activities of P. pulmonarius SMS allow its direct application to remove polycyclic aromatic hydrocarbons from contaminated soil samples [Citation165]. However, rather than directly using the bulk SMS, many applications require using isolated enzymes that can be recovered from SMS.
shows an overview of studies on the recovery of extracellular enzymes from SMS of various fungal species. The spent substrates of A. bisporus and oyster mushrooms (Pleurotus spp.) are commonly reported as sources of extracellular enzymes. Xylanases and cellulases are the most common hydrolases in the recovered enzymes, while laccases are the main reported oxidoreductases. Most studies provide a relatively detailed description of the extraction process used, while purification protocols, e.g. dialysis, ultra-filtration, anion-exchange chromatography, or gel filtration, of the extracted enzymes are not always described in detail. Production of crude enzyme extracts and their application in areas where expensive purification can be avoided is often reported [Citation164,Citation181,Citation182]. Some studies provide the exact identification of the extracted enzymes, including the complete EC classification number, while other provide trivial names or a more general classification without stating details.
Table 5. Recovery of enzymes from spent mushroom substrate. Abbrieviations used: PEG, polyethylene glycol; MnP, manganese peroxidase; LiP, lignin peroxidase; VP, versatile peroxidase.
Recovery of extracellular enzymes from SMS was reported for the first time by Ball and Jackson in 1995 using A. bisporus spent compost [Citation166]. In that study, it was found that lignocellulose-degrading enzymes can be recovered from spent mushroom compost by extraction with distilled water [Citation166]. The evaluation of the enzyme activities revealed high levels of hemicellulases (endoxylanase, β-xylosidase, xylan-acetylesterase, and arabinofuranosidase), cellulose-degrading enzymes (endoglucanase, cellobiohydrolase, and β-glucosidase), and ligninolytic enzymes (peroxidase and phenoloxidase). The activity and stability of the enzymes suggested their potential for the biological upgrading of wheat straw. After Ball and Jackson’s pioneering study, A. bisporus SMS has been studied frequently to recover enzymes by extraction with different solvent systems [Citation167–169]. Trejo-Hernandez et al. [Citation167] reported laccase extraction with Tris – HCl buffer, while Mayolo-Deloisa et al. [Citation168] developed a protocol using an aqueous potassium phosphate-polyethylene glycol two-phase system for purification of laccase extracted from A. bisporus SMS. Devi et al. [Citation169] recently reported the recovery of oxidative and hydrolytic enzymes by suspending A. bisporus SMS in sodium citrate buffer, followed by acetone precipitation and subsequent chromatographic purification. The partially purified enzyme extract was evaluated on hydrolysis of SMS polysaccharides for ethanol production.
Recovery of enzymes from SMS resulting from the cultivation of mushrooms of the genus Pleurotus has been well investigated. The first studies were published in the early 2000s, when different solvent systems, including water, sodium citrate buffer, and sodium phosphate buffer, were evaluated to extract hydrolases and oxidoreductases from SMS of P. sajor-caju SMS [Citation170] and P. ostreatus [Citation171], the latter also included SMS of other species. In other studies, different buffers and conditions were evaluated for extraction of α-amylase, endoglucanase, laccase, and endoxylanase from SMS of P. eryngii, P. ostreatus, and P. cornucopiae; the best recoveries were achieved using sodium citrate buffer [Citation172,Citation173]. Sadiq et al. [Citation174] used a sodium tartrate buffer for extracting manganese peroxidase (MnP), laccase, and lignin peroxidase (LiP) from P. ostreatus SMS and used the extract for bioremediation of contaminated soil. The SMS of P. florida [Citation36,Citation44] has also been reported as a source of lignin oxidases (versatile peroxidase (VP), MnP, LiP, and laccase) and polysaccharide hydrolases (CMCase, xylanase, and cellobiohydrolase). Crude extracts of P. pulmonarius SMS demonstrated laccase and MnP activity [Citation163]. P. pulmonarius SMS was also used to extract several hydrolases and oxidoreductases, and the extract was used for the hydrolysis of palm oil mill effluent to produce biohydrogen [Citation176]. Extraction of xylanases from P. ostreatus and P. citrinopileatus SMS has also been investigated [Citation177].
The potential for enzyme recovery from SMS of other mushroom species has also been investigated. For example, Schimpf and Schultz (2016) screened selected enzyme activities in SMSs of L. edodes, He. erinaceus, Stropharia rugosoannulata, Fomes fomentarius, and Gr. frondosa and developed a protocol for recovery of lignocellulolytic enzymes from L. edodes SMS [Citation178]. In another study, SMS from the cultivation of Au. auricula-judae, Coprinus comatus, Agrocybe cylindracea, He. erinaceus, and H. marmoreus have also been investigated as a source of xylanases [Citation177]. Screening of enzymes extracted from SMS of L. edodes, H. marmoreus, F. velutipes, and three Pleurotus strains revealed higher activity of cellulose-degrading enzymes for L. edodes extract, while the extracts of Pleurotus strains displayed higher laccase activity and ability to decolorize Coomassie Brilliant Blue [Citation179].
Recovery of laccase and several hydrolases from L. edodes, P. ostreatus, He. erinaceus, F. velutipes SMS has also been reported [Citation171].
Enzyme preparations with high xylanase activity were obtained from extracts from Tremella fuciformis SMS purified by ammonium sulfate precipitation and gel filtration chromatography [Citation177]. The purified enzyme showed good thermal stability and potential for saccharification of xylan contained in wheat bran, sugarcane bagasse, and other biomass residues. Optimal conditions for laccase extraction from G. lucidum SMS and utilization of the extract to remove toxic chemicals from an aqueous environment have also been reported [Citation180].
5.1.2. Using SMS as substrate for cultivation of enzyme-producing organisms
Since SMS is rich in nutrients and contains potential carbon sources, it can be used as a substrate for producing enzymes by cultivating enzyme-producing organisms. SMS has been used to cultivate fungi of the genus Trichoderma, the most relevant for industrial production of cellulases [Citation183]. Production of cellulases requires a cellulosic substrate for inducing the enzyme system Trichoderma spp., which consists of endoglucanases, exoglucanases, and β-glucosidases [Citation184]. Cellulose contained in lignocellulosic materials is a more suitable inducer than other alternatives, which are too expensive for industrial-scale use. Before cultivation of a cellulase producer, lignocellulose requires being pretreated, for example by a hydrothermal process [Citation185], to remove lignin and facilitate enzyme access to cellulose. A drawback of hydrothermal pretreatment is that it leads to the formation of by-products, such as furan aldehydes, aliphatic acids, and phenolic compounds, which are inhibitors of microorganisms and enzymes [Citation186]. Using SMS avoids the downsides of pretreatment since – during mushroom cultivation – lignin and part of the polysaccharides are degraded without forming inhibitors [Citation187], and, therefore, the substrate is prepared for being used in microbial fermentations.
Enzyme production by microorganisms cultivated on SMS has been less investigated than the extraction of enzymes from SMS not subjected to a new cultivation cycle. Pleurotus spp. SMS are among the most common ones to produce enzymes by other organisms (). Some studies report using SMS as substrates for conventional enzyme producers, while in other studies, the enzymes are produced by edible mushrooms cultivated on SMS.
Table 6. Production of enzymes by fungal cultivation on spent mushroom substrate. Abbreviations used: SSF, solid-state fermentation; SmF, submerged fermentation.
Trichoderma spp. are among the most common conventional enzyme producers cultivated on SMS, but there are also some reports on Aspergillus and Penicillium spp. In a recent study, He et al. [Citation188] reported the production of cellulases by T. reesei grown on corn cobs-based SMS from Au. polytricha, Auricularia nigricans, and P. ostreatus. In that study, cellulase production was more effective when using earlier ‘flushes’ of SMS than when several harvests were produced on the same substrate. The highest cellulase activity was obtained using the third flush of mushrooms of Au. polytricha SMS, particularly when the fermentation process was assisted with ultrasound. The study showed that higher cellulase activity could be obtained by cultivation on untreated SMS than on acid- or alkali-treated SMS. The potential of spent mushroom compost (SMC) of A. bisporus for cultivation of enzyme-producing fungi has also been shown. Production of endoglucanase, endoxylanase, and β-glucosidase using Trichoderma isolates and a strain of Aspergillus niger on A. bisporus SMC without nutrient supplementation was reported [Citation189]. SMS resulting from growing P. sajor-caju on sugarcane bagasse was used to produce cellulases and xylanases by Penicillium echinulatum [Citation190].
The production of enzymes by cultivating edible mushrooms on SMS has also been reported. P. ostreatus SMS supplemented with wheat bran and soybean flour was a suitable substrate for the cultivation of P. ostreatus, P. pulmonarius, Ganoderma adspersum, Ganoderma resinaceum, and L. edodes for producing laccase [Citation191]. The study showed good potential of the supplemented SMS for laccase production by Ganoderma spp. and fruitbodies by Pleurotus spp. In another study by the same group, laccase was produced by cultivating P. ostreatus and P. pulmonarius on P. ostreatus SMS, and the crude enzyme’s potential for removing phenolic compounds from olive mill and winery wastewaters was evaluated [Citation26].
Another approach is cultivating enzyme producers in SMS that has already been subjected to the extraction of extracellular enzymes. This approach has been tested for P. pulmonarius SMS, which was first subjected to extraction of lignin-degrading enzymes, and then used as substrate for producing cellulases by Trichoderma asperellum cultivation [Citation163]. It was also applied for P. florida SMS, which was first used as a source of laccase and several hydrolases, and then directed to the production of cellulases by either Trichoderma longibrachiatum and Aspergillus aculeatus [Citation175]. T. longibrachiatum resulted in higher activity of endoglucanase, exoglucanase, and xylanase, while As. aculeatus was a better cellobiase producer ().
5.2. Bioactive compounds
SMS contains bioactive compounds of different functionality and origin. The fungal mycelium contains polysaccharides, sterols, proteins, polyphenols, vitamins, and other bioactive molecules. Mycelial growth throughout the surrounding environment also results in the secretion of potentially useful bioactive compounds. In addition, the extractive fraction of the lignocellulosic substrate and the oligomeric products from fungal degradation of polysaccharides and lignin might also be sources of bioactive substances. However, while the bioactive molecules of the sporocarps of edible fungi have been extensively investigated [Citation192], the information on the bioactive potential available in SMS is still limited. Recovery of bioactive compounds is a promising direction for valorizing SMS.
5.2.1. Polysaccharides
Polysaccharides are among the bioactive substances responsible for the immunomodulatory and antitumor effects of edible and medicinal mushrooms [Citation193]. However, in most of the research dealing with fungal polysaccharides the investigated sources are fruitbodies or mycelia [Citation194], while extraction from SMS has been less explored. A study on extraction and characterization of a polysaccharide from L. edodes SMS published in 2012 by Zhu et al. provided the start for the research on SMS as a source of bioactive molecules [Citation195]. Henceforth, several relevant reports on obtaining bioactive extracts from SMS have been published. Most publications deal with L. edodes, Pleurotus spp., and Ganoderma spp., but SMS from the cultivation of other fungal species has also been investigated ().
Table 7. Recovery of bioactive compounds from spent mushroom substrate. Abbreviations used: UAE, ultrasound-assisted extraction; SSF, solid-state fermentation.
Water extraction at temperatures around 80–90°C, followed by alcohol precipitation, and chromatographic purification, is a standard procedure for recovering polysaccharides from SMS. Accordingly, a heteropolysaccharide displaying antibacterial activity against three different microorganisms was recovered from L. edodes SMS [Citation195]. The same method, combined with partial hydrolysis, either chemical [Citation57] or enzymatic [Citation196], was applied to L. edodes SMS for extracting polysaccharides showing antioxidant, anti-inflammatory, and renoprotective activities. Water extraction has also been reported to extract polysaccharides from G. lucidum SMS [Citation197], and to extract β-glucans and other compounds from rice husk-based SMS of Pycnoporus sanguineus and Panus strigellus (syn. Pleurotus tubarius) [Citation198].
Extraction with aqueous alkaline solutions is another useful method for recovering polysaccharides. He et al. [Citation199] reported the obtention of a polysaccharide extract from P. eryngii SMS by alkaline extraction followed by deproteinization and gel filtration chromatography. The refined product was a polysaccharide-protein complex containing 99% (w/w) of a polysaccharide composed of anhydroxylose, anhydroglucose, and anhydroarabinose units. Strong antioxidant activity – with potential food applications – was revealed in vitro for the polysaccharide-protein complex. A comparable extraction approach has also been used to recover polysaccharides from L. edodes SMS [Citation193]. Exhaustive characterization revealed that the L. edodes SMS extract contained heteropolysaccharides exerting antiproliferative effects against six tested human tumor cell lines.
Subcritical water extraction (SWE) can be applied to extract bioactive molecules. SWE of polysaccharides from P. ostreatus SMS and L. edodes residual basidiocarps by autoclaving at 120°C has been reported [Citation139].
Partial enzymatic hydrolysis can also be used to extract polysaccharides from SMS. Hydrolysis with cellulases for two hours was used for recovering polysaccharides from the SMS of C. militaris [Citation200]. Four polysaccharide fractions were isolated, and three displayed good antioxidant activity with no cytotoxicity. Enzyme treatment has also been used to recover polysaccharides from SMS of Ag. cylindracea, L. edodes, H. marmoreus, P. ostreatus and C. militaris [Citation201]. The polysaccharides were isolated from the extracts by ethanol precipitation and purified by deproteinization with the Sevag regent, and their antioxidant activity was evaluated in vitro. The polysaccharides from Ag. cylindracea SMS had the best oxygen free radical-scavenging capacity and ferric reducing power (FRAP), while those from Hy. marmoreus and P. ostreatus displayed the best ABTS and DPPH radical scavenging activities.
Another way of producing chemical compounds of interest is to use SMS as a substrate for cultivating other organisms. For instance, P. ostreatus SMS was reported to be used for producing crude exopolysaccharides by cultivation of P. ostreatus and P. pulmonarius [Citation191].
5.2.2. Sterols and other compounds
Ergosterol, the most abundant sterol in fungi, has relevant biological activities for food, pharmaceutical, and biomedical uses, and it is a precursor of vitamin D2. Most of the reports on ergosterol extraction from mushroom residues deal mainly with stipes of fruitbodies or mushrooms not meeting commercial specifications [Citation203]. However, the potential of L. edodes SMS as a source of ergosterol has recently been demonstrated [Citation58]. Ergosterol-rich extracts were obtained from L. edodes SMS using ultrasound-assisted extraction, a non-conventional technique for extracting natural products from various biomaterials; in vitro experiments revealed that the produced extracts have antitumor activity against three cancer cell lines.
The presence of steroids and saturated terpenes has been shown in water extracts of SMS from the cultivation of Py. sanguineus and Pa. strigellus (syn. P. tubarius) SMS on rice husk [Citation198]. The purine analog pentostatin, a potent anticancer drug, was produced by cultivating a cellulose-degrading transformant of C. militaris using H. marmoreus SMS as substrate [Citation202].
Phenolic compounds can also be extracted from SMS. Elsakhawy et al. reported the production of phenol-rich extracts from P. ostreatus SMS using either 0.5 N NaOH [Citation152] or tap water [Citation153] as solvents. The produced extracts were further assayed as plant-disease control and biofertilizer.
6. Spent mushroom substrate valorization as part of cascade use of plant biomass
The generation of plant biomass resources by agriculture and forestry takes a long time and requires considerable land areas; thus, their utilization should be rational and efficient. Residual biomass materials, such as side/waste streams or byproducts from varying stages of production/processing chains, contain components of high potential for value-added applications. A common approach for biomass valorization today is to burn it in a resource-inefficient way to generate heat and power for energy purposes. For bioeconomy development in a resource-efficient way, cascade use of plant biomass should always be considered. Cascade use, also known as cascading use [Citation204], is a complex interaction of material flows used as a strategy to increase resource efficiency in biomass processing. Cascade use occurs when biomass is processed through a series of material uses (), by reuse and recycling, before finally being used for energy recovery [Citation205].
Figure 3. Schematic illustration of the cascade-use concept of bioresources (based on Vis et al. 2016 [Citation204], p. 6, https://data.europa.eu/doi/10.2873/827106).
![Figure 3. Schematic illustration of the cascade-use concept of bioresources (based on Vis et al. 2016 [Citation204], p. 6, https://data.europa.eu/doi/10.2873/827106).](/cms/asset/eb3cdc96-9aa6-4b92-8462-f5f166cd8e84/kbie_a_2252138_f0003_oc.jpg)
Cultivation of edible mushrooms plays a unique role in supplying highly nutritive and health-promoting food. Still, it generates vast amounts of SMS, which is mostly discarded or inefficiently used despite its potential for generating value-added products. As a cellulose-rich bioresource, SMS can also be considered a material of interest for developing sugar-platform applications after enzymatic saccharification. Compared with the lignocellulosic materials used to formulate the initial mushroom substrate, SMS is more susceptible to biochemical conversion using enzymes and microorganisms [Citation16,Citation187,Citation206]. That is mainly because the cultivation of white-rot edible fungi constitutes a biological process that modifies lignocellulose by removing a large part of lignin and hemicelluloses, which interferes with the enzymatic saccharification of cellulose. Furthermore, SMS, i.e. a material resulting from the aforementioned process (which could be considered as a biological pretreatment), contains few inhibitory compounds or external chemicals that might negatively affect downstream processing or harm the environment [Citation187,Citation206].
Applying cascade uses to processing plant biomass by mushroom cultivation combined with SMS valorization through biochemical conversion and other approaches, is expected to maximize the cost-effectiveness of a value chain of variable potential products. The cascade-use concept also results in minimizing resource loss and environmental impacts. Following a cascading approach, SMS, as the primary by-product of mushroom cultivation can be re-used as raw material for new processes, extending total biomass availability within the system. That is a rational approach, where different valuable biomass constituents are recovered and converted into value-added products. Energy uses of residual biomass are considered only at the end of the life cycle when all higher-value products and services have been exhausted. There are different possible examples of multi-stage cascading uses for SMS valorization. Three promising published case studies are discussed in this section.
6.1. Case study 1: food – ethanol – solid fuel
The integrated production of L. edodes mushroom (food) and biofuels from hardwood residues can be an example of cascade use [Citation16,Citation187,Citation206,Citation207]. Food (mushrooms) is produced on a lignocellulosic substrate. Concomitantly, mushroom cultivation selectively degrades lignin and hemicelluloses, thus facilitating the enzymatic saccharification of cellulose. Glucose from the saccharification process can then be fermented to ethanol using yeast. Enzymatic saccharification also generates lignin-rich solid leftovers, which can be used as a solid fuel ().
Figure 4. Schematic illustration of the concept food – ethanol – solid fuel. The figure is modified from Chen, 2021 [Citation208], p. 15, https://pub.epsilon.slu.se/26324/1/chen_f_211216.pdf.
![Figure 4. Schematic illustration of the concept food – ethanol – solid fuel. The figure is modified from Chen, 2021 [Citation208], p. 15, https://pub.epsilon.slu.se/26324/1/chen_f_211216.pdf.](/cms/asset/1b8a87a9-607a-4f78-8895-2d98c54fc497/kbie_a_2252138_f0004_oc.jpg)
From a circular bioeconomy point of view, the forest residues can be considered primary bioresources from forest production (c.f. ). Exploiting the forest residues as mushroom growing substrates results in the production of fruitbodies as primary products. SMS is the secondary bioresource and can be converted to the secondary product ethanol. Ethanol can be used to synthesize renewable polyethylene to produce green plastics or for fuel applications, including advanced jet biofuels. After cellulose saccharification, the solid leftover can also be recovered as a tertiary bioresource/biowaste, and converted to solid fuel, a ‘tertiary product.’ A recently published mass balance analysis revealed that one ton of birch-based initial mushroom substrate might result in about 600 kg of fresh shiitake (L. edodes) fruitbodies (90% moisture), 130 liter of ethanol, and 300 kg (dry mass) of solid biofuel [Citation187]. This system/approach can also be applied to other mushroom species. Using an experimental setting like the one used for L. edodes, Chen et al. [Citation209] found that one ton of birch-based initial substrate might result in about 400 kg of fresh fruitbodies (90% moisture) of Au. auricula-judae, 35 liters of ethanol, and 300 kg dry mass of solid biofuel. The solid fuel was found to have a relatively high calorific value and favorable characteristics for direct combustion to produce heat. The generated heat can be used for the pasteurization of substrates or space heating.
The outcome of the production chain can be affected by the composition of the initial substrates used for mushroom cultivation. Chen et al. [Citation206] reported that alder-based substrate led to 4% more mushroom fruitbodies, 14% more ethanol, and 23% more solid fuel than birch-based substrate. On the other hand, an aspen-based substrate resulted in a 37% lower yield of fruitbodies than the birch-based one, although the yields of ethanol and solid fuel were comparable for substrates from both tree species.
The same concept is also applicable to other biofuels. For instance, another ‘food – biofuel – solid fuel’ alternative is to produce biogas instead of ethanol as a secondary product. Lin et al. [Citation210] cultivated shiitake on woodchips and produced biogas by anaerobic digestion (AD) of SMS; at the end, around 53–57% (dry mass) of the substrate was solid leftover. Since the AD process consumes mostly carbohydrates [Citation211,Citation212], the leftovers are expected to have a relatively high content of lignin, a component with a high calorific value. Therefore, using the leftovers as a solid fuel for a self-supporting heating system could be meaningful. In a slightly different alternative, rice straw was used as the main ingredient for P. ostreatus substrate, fruitbodies were produced as the primary product, SMS was directed to AD for producing biomethane as a secondary product, and the AD digestate was used as biofertilizer (tertiary product) for rice cultivation [Citation213].
6.2. Case study 2: food – biogas − 2nd cycle mushroom
Another example of a cascade system was reported by Ikeda et al. [Citation214] using the mushroom ‘enokitake’ (F. velutipes) cultivated in a substrate based on corncobs supplemented with rice bran. In this case, agricultural residues were the primary bioresource, and enokitake fruitbodies were the primary product. The SMS resulting after mushroom harvest was anaerobically digested for producing biogas, the secondary product. After the AD process, around 45% of the initial mass was left as solid residue or digestate. In the next step, KOH or NaOH was used for pretreating the AD residue (SMS-ADR), which was then mixed with rice bran at a 50:50 weight ratio for formulating a new substrate to be subsequently used for a second enokitake cultivation cycle. The results were promising: the yield of the tertiary product, i.e. mushrooms cultivated on SMS-ADR, was comparable to those of the primary one, i.e. mushrooms cultivated on corncob-based ‘standard’ substrate. Crude protein, ether-extracted compounds, crude fiber, minerals (Na, P, Ca, K, Mg), and free amino acids in fruitbodies showed similar content to those obtained from the standard substrates. The study did not discuss further use of the second cycle SMS (SMS-II). In our opinion, this cascading system could still be extended to a quaternary product by exploiting the potential of SMS-II as a biofertilizer or a soil amendment.
Cascade systems including biogas as the secondary product are also feasible for other mushroom species. Since lignin content decreases during the mushroom cultivation, the resulting SMS is accessible to anaerobic microbes, thus facilitating AD conversion of carbohydrates to biogas. On the other hand, after biogas production, and although data on the chemical composition of the digestate are not available [Citation215], the lignin content is expected to increase due to its recalcitrance to degradation by anaerobic bacteria [Citation211,Citation212,Citation216]. Therefore, it was wise to choose white-rot fungi again to break the recalcitrance of lignin to produce additional value-added tertiary products. Although the low pH and the presence of unknown by-products may inhibit a second mushroom cultivation cycle, it was shown that KOH or NaOH soaking was a viable method to improve the susceptibility of digestate to be further used for enokitake cultivation [Citation214]. Nevertheless, the precise mechanism behind the alkaline reactivation of the digestate from AD for mushroom cultivation remains to be clarified.
6.3. Case study 3: food − 2nd cycle mushroom – enzymes
Another possible cascading chain can include two cycles of mushroom cultivation in a row followed by recovery of extracellular enzymes as a tertiary product. Economou et al. [Citation191] reported a case study, where oyster mushroom (P. ostreatus) was produced on a wheat straw-based substrate, and the resulting SMS (SMS-I) was tested as the main ingredient of the substrate for a second mushroom cultivation cycle. After harvesting the fruitbodies from the second production cycle, the generated SMS (SMS-II) was used as a source for the recovery of the lignin-degrading enzyme laccase. Among the five fungal species tested for the second mushroom production cycle, P. pulmonarius resulted in the SMS providing the highest yield of laccase, 2465 U g−1 day−1 (dry mass based). The crude laccase extract was then used for the dephenolization of wastewaters [Citation26]. The authors did not explore possible uses of the solid stream remaining after laccase extraction from SMS-II. A potential extension of the cascading system would be possible by using that stream as either biofertilizer or solid fuel.
Using P. ostreatus as the species involved in the first step of the cascading system is a reasonable strategy considering that Pleurotus spp. are among the most studied white-rot fungi for biological treatments of lignocellulosic materials [Citation217]. Compared with other edible fungi, they have the advantages of presenting a relatively shorter life cycle, and broader adaptation to substrate assortments and growing environments. Even though Pleurotus lignocellulolytic enzyme activities are generally comparable to those of L. edodes [Citation215], the lignocellulose degradation capacity of the former is generally lower than that of shiitake, probably because of the shorter life cycle [Citation217]. It must be emphasized that the determination of SMS composition, which is essential for fully understanding the potential of the SMS to be directed to new cultivation cycles, is often underestimated in the literature.
The cultivation of white-rot edible fungi on primary bioresources results to food (mushrooms), and functions as a biological pretreatment for facilitating biochemical conversions. Therefore, mushroom cultivation is crucial in cascading systems of lignocellulosic biomass utilization. In addition to the case studies discussed above, many other cascade systems producing fruitbodies as a primary product, and including other products or services, can be proposed to valorize SMS and other residual streams. The feasibility of producing antibiotics [Citation218], antitumor sterols [Citation58], seedbed of vegetables [Citation219], fertilizers [Citation104], soil bioremediation agents [Citation220], enzymes [Citation175], biochar [Citation221], and other products has been demonstrated. Some products could be considered as different ‘puzzle pieces’ to be chosen and integrated into a chain of cascade uses. However, appropriate approaches ensuring optimal process integration remain to be developed. Process integration has to be developed through interdisciplinary approaches to maximize system values for the circular bioeconomy and the protection of the environment. Furthermore, systematic evaluation (e.g. life cycle assessment) and optimization of processes for the cascading uses must be addressed.
7. Future directions and conclusions
Cultivation of edible and medicinal mushrooms is a very dynamic business, with an impressing development during the last decades. However, increased mushroom production leads to the generation of high quantities of spent mushroom substrate (SMS). The accumulation of non-used SMS, or its limited or not high-added value applications, undermines the future of pertinent commercial activities. Therefore, achieving an efficient valorization of SMS – beyond its current low-value use – is of paramount importance for the sustainable development of the mushroom industry. The research discussed in this review shows the vast potential of SMS as a source of valuable products and services.
The presence of valuable nutrients and energy sources for supporting new cultivation processes make SMS a suitable substrate component for new mushroom cultivation cycles provided that a suitable treatment or supplementation is applied. Reusing SMS in new cultivation of mushrooms of either the same or other species has significant potential for reaching high yields in an environmentally sustainable way and at the same time contribute to the reduction of production costs. The high nutritional value of SMS could also be exploited for the development of new feeds; the output of recent experimental work convincingly shows the feasibility of including SMS in the diets of poultry, ruminants, and monogastric animals, as well as beyond traditional husbandry, in pisciculture and insect farming. However, making SMS a regular diet ingredient poses complex challenges related to its fiber content and digestibility, and the acceptance by the animal. Recent research has faced those downsides since it has been demonstrated that by application of appropriate treatments the nutritional and acceptance features of SMS can be enhanced.
SMS’s physical properties and chemical composition support the development of novel environment-friendly and cost-effective bio-based products, which can be used as part of sustainable agronomic practices in substituting fossil-based fertilizers and synthetic pesticides. Well-designed experiments have shown that SMS application as a soil amendment or fertilizer has beneficial effects on soil fertility and structure, without causing secondary salinization or acidification. The reported research also shows the potential of SMS as source of products for biological control against plant diseases, and its favorable effect on the production of secondary metabolites in plants and at enhancing the nutritional value of the fruits and vegetables. Scaling up the experimentation to large-scale greenhouse and field trials is required. Furthermore, increasing demonstration actions are expected to fully demonstrate/evidence the potential of SMS within a sustainable agriculture model.
SMS contains extracellular enzymes secreted during fungal growth and used to degrade the substrate’s macromolecules. Those enzymes make it possible to use the SMS in services such as decolorization of textile effluents, bioremediation of contaminated soil, and wastewater treatment. Enzymes can also be extracted from SMS using different solvent systems. Furthermore, the potential of SMS as substrate for the cultivation of enzyme-producing microorganisms has been shown. The crude extracts of SMS enzymes can be subjected to various degrees of purification rendering refined preparations suitable for added-value applications, where enzyme purity is a decisive criterion.
Several publications report on exploiting SMS bioactive compounds for various uses. However, the extraction of bioactive compounds from SMS is just an emerging area of high interest. Recently published results indicated that SMS bioactive molecules could be used as added-value, sustainable, bio-based ingredients in socially-sensitive business sectors. SMS-derived nutraceuticals, food supplements, functional foods, and active ingredients might be the foundation of a new ‘next-generation mycotherapeuticals’ sector. That would require developing appropriate protocols for extracting bioactive molecules from SMS, a task that faces major challenges regarding the extraction’s effectiveness without affecting the properties of the molecules of interest, and by avoiding the degradation of non-targeted compounds that might also be of interest.
Applying the cascade-use concept to SMS valorization is essential to increase resource efficiency in biomass processing and mushroom production. Arranging different alternatives of SMS utilization in cascading systems, where mushroom production is included as the primary process and the by-products are converted to value-added products, will result in a value chain with minimal resource losses and with no adverse environmental impact, in agreement with the principles of sustainable development. Appropriate implementation of the cascade-use concept requires significant efforts to ensure optimal process integration based on interdisciplinary approaches. By achieving it, the system values can be maximized, and the extensive use of SMS for generating high-value products and services within a circular bioeconomy scenario can become a reality.
Author contributions statement
CM, GIZ, and SX conceived the presented idea, developed the methodology, wrote sections of the original draft, reviewed and edited the whole manuscript, and contributed to the visualization. GK wrote sections of the original draft, reviewed and edited the whole manuscript, and contributed to the visualization. KOS reviewed and edited the whole manuscript and contributed to the visualization.
Ethics statement
This study was conducted in an ethical and responsible manner and is in full compliance with all relevant codes and legislation. No experiments on humans or animals were performed. The work does not involve biological material, protected or nonpublic datasets, collections or sites.
Acknowledgments
Sarah Jasmin Klausen (Inland Norway University of Applied Sciences) is thanked for the graphic design.
Disclosure statement
One of the authors (C.M.) is a member of the Editorial Board. The authors declare that they do not have other potential conflicts of interest.
Data availability statement
Since no new data were created by the authors, data sharing is not applicable regarding their own research results. Raw data on search criteria used for building the review, as well as the excluded material, will be available upon reasonable request.
Additional information
Funding
References
- Maurice S, Arnault G, Nordén J, et al. Fungal sporocarps house diverse and host-specific communities of fungicolous fungi. Isme J. 2021;15(5):1445–40. doi: 10.1038/s41396-020-00862-1
- El-Ramady H, Abdalla N, Badgar K, et al. Edible mushrooms for sustainable and healthy human food: nutritional and medicinal attributes. Sustainability. 2022;14(9):4941. doi: 10.3390/su14094941
- Gargano ML, van Griensven LJLD, Isikhuemhen OS, et al. Medicinal mushrooms: Valuable biological resources of high exploitation potential. Plant Biosyst - An Int J Dealing Aspects Plant Biol. 2017;151(3):548–565. doi: 10.1080/11263504.2017.1301590
- Carrasco-González JA, Serna-Saldívar SO, Gutiérrez-Uribe JA. Nutritional composition and nutraceutical properties of the Pleurotus fruiting bodies: Potential use as food ingredient. J Food Compos Anal. 2017;58:69–81. doi: 10.1016/j.jfca.2017.01.016
- de Frutos P. Changes in world patterns of wild edible mushrooms use measured through international trade flows. For Policy Econ. 2020;112:102093. doi: 10.1016/j.forpol.2020.102093
- Royse DJ, Baars J, Tan Q. Current overview of mushroom production in the world. In: Edible and medicinal mushrooms. John Wiley & Sons, Ltd; 2017. pp. 5–13. doi: 10.1002/9781119149446.ch2
- Kumla J, Suwannarach N, Sujarit K, et al. Cultivation of mushrooms and their lignocellulolytic enzyme production through the utilization of agro-industrial waste. Molecules. 2020;25(12):2811. doi: 10.3390/molecules25122811
- Kaushal LA, Prashar A. Agricultural crop residue burning and its environmental impacts and potential causes – case of northwest India. J Environ Plan Manag. 2021;64(3):464–484. doi: 10.1080/09640568.2020.1767044
- Martín C. Pretreatment of crop residues for bioconversion. Agronomy. 2021;11(5):924. doi: 10.3390/agronomy11050924
- Chen L, Qian L, Zhang X, et al. Research progress on indoor environment of mushroom factory. Int J Agric Biol Eng. 2022;15(1):25–32. doi: 10.25165/j.ijabe.20221501.6872
- Zisopoulos FK, Ramírez HAB, Goot AJ, et al. A resource efficiency assessment of the industrial mushroom production chain: the influence of data variability. J Clean Prod. 2016;126:394–408. doi: 10.1016/j.jclepro.2016.03.066
- Atallah E, Zeaiter J, Ahmad MN, et al. Hydrothermal carbonization of spent mushroom compost waste compared against torrefaction and pyrolysis. Fuel Process Technol. 2021;216:106795. doi: 10.1016/j.fuproc.2021.106795
- Beyer DM Impact of the mushroom industry on the environment [Internet]. Penn State Extension. Pennsylvania State University; 2011 [cited 2022 Jul 18]. Available from: https://extension.psu.edu/impact-of-the-mushroom-industry-on-the-environment
- Council directive 1999/31/EC on the landfill of waste [Internet]. 1999. Available from: http://ec.europa.eu/environment/waste/landfill_index.htm.
- Gong F, Zhang D, Zgang Y, et al. The nutritional value and safety assessment analysis on spent mushroom substrate of Pleurotus ostreatus. China Anim Husb Vet Med. 2012;39(11):86.
- Chen F, Martín C, Finell M, et al. Enabling efficient bioconversion of birch biomass by Lentinula edodes: regulatory roles of nitrogen and bark additions on mushroom production and cellulose saccharification. Biomass Convers Biorefin. 2022;12(4):1217–1227. doi: 10.1007/s13399-020-00794-y
- Chen F, Xiong S, Sundelin J, et al. Potential for combined production of food and biofuel: Cultivation of Pleurotus pulmonarius on soft- and hardwood sawdusts. J Clean Prod. 2020;266:122011. doi: 10.1016/j.jclepro.2020.122011
- Wei M, Geladi P, Xiong S. NIR hyperspectral imaging and multivariate image analysis to characterize spent mushroom substrate: a preliminary study. Anal Bioanal Chem. 2017;409(9):2449–2460. doi: 10.1007/s00216-017-0192-2
- Mohd Hanafi FH, Rezania S, Mat Taib S, et al. Environmentally sustainable applications of agro-based spent mushroom substrate (SMS): an overview. J Mater Cycles Waste Manag. 2018;20(3):1383–1396. doi: 10.1007/s10163-018-0739-0
- Leong YK, Ma TW, Chang JS, et al. Recent advances and future directions on the valorization of spent mushroom substrate (SMS): A review. Bioresour Technol. 2022;344:126157. doi: 10.1016/j.biortech.2021.126157
- Koutrotsios G, Kalogeropoulos N, Kaliora AC, et al. Toward an increased functionality in oyster (Pleurotus) mushrooms produced on grape marc or olive mill wastes serving as sources of bioactive compounds. J Agric Food Chem. 2018;66(24):5971–5983. doi: 10.1021/acs.jafc.8b01532
- Koutrotsios G, Tagkouli D, Bekiaris G, et al. Enhancing the nutritional and functional properties of Pleurotus citrinopileatus mushrooms through the exploitation of winery and olive mill wastes. Food Chem. 2022;370:131022. doi: 10.1016/j.foodchem.2021.131022
- Zervakis GI, Koutrotsios G. Solid-state fermentation of plant residues and agro-industrial wastes for the production of medicinal mushrooms. In: Agrawal D, Tsay H, Shyur L, Wu Y Wang S, editors. Medicinal plants and fungi: Recent advances in research and development. Singapore: Springer Singapore; 2017. pp. 365–396. doi: 10.1007/978-981-10-5978-0_12
- Vega A, De León JA, Miranda S, et al. Agro-industrial waste improves the nutritional and antioxidant profile of Pleurotus djamor. Clean Waste Syst. 2022;2:100018. doi: 10.1016/j.clwas.2022.100018
- Cunha Zied D, Sánchez JE, Noble R, et al. Use of spent mushroom substrate in new mushroom crops to promote the transition towards a circular economy. Agronomy. 2020;10(9):1239. doi: 10.3390/agronomy10091239
- Economou CN, Philippoussis AN, Diamantopoulou PA. Spent mushroom substrate for a second cultivation cycle of Pleurotus mushrooms and dephenolization of agro-industrial wastewaters. FEMS Microbiol Lett. 2020;367(8). doi: 10.1093/femsle/fnaa060
- Mamiro DP, Royse DJ, Beelman RB. Yield, size, and mushroom solids content of Agaricus bisporus produced on non-composted substrate and spent mushroom compost. World J Microbiol Biotechnol. 2007;23(9):1289–1296. doi: 10.1007/s11274-007-9364-0
- Wu CY, Liang CH, Liang ZC. Evaluation of using spent mushroom sawdust wastes for cultivation of Auricularia polytricha. Agronomy. 2020;10(12):1892. doi: 10.3390/agronomy10121892
- Ahmad Zakil F, Mohd Isa R, Mohd Sueb MS, et al. Growth performance and mineral analysis of Pleurotus ostreatus (oyster mushroom) cultivated on spent mushroom medium mixed with rubber tree sawdust. Mater Today Proc. 2022;57:1329–1337. doi: 10.1016/j.matpr.2022.01.112
- Lisiecka J, Prasad R, Jasinska A. The utilisation of Pholiota nameko, Hypsizygus marmoreus, and Hericium erinaceus spent mushroom substrates in Pleurotus ostreatus cultivation. Horticulturae. 2021;7(10):396. doi: 10.3390/horticulturae7100396
- Lin Q, Long L, Wu L, et al. Evaluation of different agricultural wastes for the production of fruiting bodies and bioactive compounds by medicinal mushroom Cordyceps militaris. J Sci Food Agric. 2017;97(10):3476–3480. doi: 10.1002/jsfa.8097
- Zeng XL, Han F, Ye JL, et al. Recycling spent Pleurotus eryngii substrate supplemented with Tenebrio molitor feces for cultivation of Agrocybe chaxingu. Int J Recycl Org Waste Agricult. 2017;6(4):275–280. doi: 10.1007/s40093-017-0171-9
- Noonsong V. Recycling of spent Pleurotus compost for production of the Agrocybe cylindracea. Mycosphere. 2016;7(1):36–43. doi: 10.5943/mycosphere/7/1/4
- Picornell-Buendía R, Pardo-Giménez A, de Juan-Valero JA. Agronomic assessment of spent substrates for mushroom cultivation. Biotechnol Agron Soc Environ. 2016;20(3):263–374. doi: 10.25518/1780-4507.13138
- Picornell MR, Pardo A, de Juan JA. Reuse of degraded Pleurotus ostreatus substrate through supplementation with wheat bran and Calprozime® quantitative parameters. Agron Colomb. 2015;33(2):261–270. doi: 10.15446/agron.colomb.v33n2.49760
- Wang S, Xu F, Li Z, et al. The spent mushroom substrates of Hypsizigus marmoreus can be an effective component for growing the oyster mushroom Pleurotus ostreatus. Sci Hortic. 2015;186:217–222. doi: 10.1016/j.scienta.2015.02.028
- Ashrafi R, Mian MH, Rahman MM, et al. Recycling of spent mushroom substrate for the production of oyster mushroom. Res Biotechnol. 2014;5:37–41.
- Mishra KK, Pal RS, Arunkumar R, et al. Antioxidant properties of different edible mushroom species and increased bioconversion efficiency of Pleurotus eryngii using locally available casing materials. Food Chem. 2013;138(2–3):1557–1563. doi: 10.1016/j.foodchem.2012.12.001
- Sun X, Xu H, Liang H, et al. Chemical composition of spent Pleurotus eryngii mushroom substrate and its reuse for Volvariella volvacea production. Asian J Chem. 2013;25(18):10504–10508. doi: 10.14233/ajchem.2013.15809
- Zhang W, Niu X, Zhang W, et al. The cultivation of Agaricus bisporus on the spent substrate of Flammulina velutipes. African Journal Of Agricultural Research. 2013;8:4860–4863.
- González Matute R, Figlas D, Curvetto N. Agaricus blazei production on non-composted substrates based on sunflower seed hulls and spent oyster mushroom substrate. World J Microbiol Biotechnol. 2011;27(6):1331–1339. doi: 10.1007/s11274-010-0582-5
- Pardo-Giménez A, Pardo-González JE, Zied DC. Evaluation of harvested mushrooms and viability of Agaricus bisporus growth using casing materials made from spent mushroom substrate. Int J Food Sci Technol. 2011;46(4):787–792. doi: 10.1111/j.1365-2621.2011.02551.x
- Gern RMM, Libardi Junior N, Patrício GN, et al. Cultivation of Agaricus blazei on Pleurotus spp. spent substrate. Braz Arch Biol Technol. 2010;53(4):939–944. doi: 10.1590/S1516-89132010000400024
- Pardo-Giménez A, Pardo-González JE. Elaboration of new substrates for cultivating Pleurotus ostreatus (Jacq.) P. Kumm. based on degraded substrates from edible fungi cultivation. ITEA. 2009;105(2):89–98.
- Singh C, Oys S. Recycling of spent oyster mushroom substrate to recover additional value. KUSET. 2009;5.
- Liang ZC, Wu C, Wang J. The evaluation of using mushroom sawdust wastes for cultivation of Pleurotus citrinopileatus. Fungal Sci. 2005;20:27–34.
- Shashirekha MN, Rajarathnam S, Bano Z. Enhancement of bioconversion efficiency and chemistry of the mushroom, Pleurotus sajor-caju (Berk and br.) Sacc. produced on spent rice straw substrate, supplemented with oil seed cakes. Food Chem. 2002;76(1):27–31. doi: 10.1016/S0308-8146(01)00244-8
- Chai J, Kim Y, Lee S. Reutilization of enokitake cultural waste as cultivating substrates for production of shiitake, Lentinus edodes and enokitake, Flammulina velutipes. Gwacheon, South Korea: Report of the Ministry of Agriculture & Forestry of the Republic of Korea; 2000. p. 113.
- Kilpatrick M, Murray DJ, Ward F Influence of substrate formulation and autoclave treatment on Lentinula edodes production. Proceedings of the 15th International Congress on the Science and Cultivation of Edible Fungi. Maastricht, Netherlands; 2000. p. 803–810.
- Nakaya M, Yoneyama S, Kato Y, et al. Cultivation of some important edible mushrooms using the sawdust from waste shiitake bed logs. Proceedings of the 3rd International Conference of Mushroom Biology and Mushroom Products. Sydney, Australia: World Society for Mushroom Biology and Mushroom Products; 1999 [cited 2022 May 20]. Available from:https://www.fungifun.org/docs/mushrooms/Shiitake/Mushworld%20Cultivation%20of%20some%20important%20edible%20mushrooms%20using%20the%20sawdust%20from%20waste%20Shiitake%20bed%20logs.pdf
- Akamatsu Y. Reutilization of culture wastes of Pleurotus ostreatus and Phollota nameko for cultivation of Lyophyllum decastes. J Wood Sci. 1998;44(5):417–420. doi: 10.1007/BF01130458
- Royse DJ. Recycling of spent shiitake substrate for production of the oyster mushroom, Pleurotus sajor-caju. Appl Microbiol Biotechnol. 1992;38(2):179–182. doi: 10.1007/BF00174464
- FAO. The future of food and agriculture – Alternative pathways to 2050 | Global Perspectives Studies | Food and Agriculture Organization of the United Nations. 2018.
- Colgrave ML, Dominik S, Tobin AB, et al. Perspectives on future protein production. J Agric Food Chem. 2021;69(50):15076–15083. doi: 10.1021/acs.jafc.1c05989
- Boontiam W, Wachirapakorn C, Wattanachai S. Growth performance and hematological changes in growing pigs treated with Cordyceps militaris spent mushroom substrate. Vet World. 2019;13(4):768–773. doi: 10.14202/vetworld.2020.768-773
- Chuang WY, Hsieh YC, Lee TT. The effects of fungal feed additives in animals: a review. Animals (Basel). 2020;10(5):805. doi: 10.3390/ani10050805
- Song X, Ren Z, Wang X, et al. Antioxidant, anti-inflammatory and renoprotective effects of acidic-hydrolytic polysaccharides by spent mushroom compost (Lentinula edodes) on LPS-induced kidney injury. Int j biol macromol. 2020;151:1267–1276. doi: 10.1016/j.ijbiomac.2019.10.173
- Wang Q, Cheng J, Wang L, et al. Valorization of spent shiitake substrate for recovery of antitumor fungal sterols by ultrasound-assisted extraction. J Food Biochem. 2018;42(5):e12602. doi: 10.1111/jfbc.12602
- Ivarsson E, Grudén M, Södergren J, et al. Use of faba bean (Vicia faba L.) hulls as substrate for Pleurotus ostreatus – potential for combined mushroom and feed production. J Clean Prod. 2021;313:127969. doi: 10.1016/j.jclepro.2021.127969
- Hsieh YC, Lin WC, Chuang WY, et al. Effects of mushroom waster medium and stalk residues on the growth performance and oxidative status in broilers. Anim Biosci. 2021 ;34(2):265–275. doi: 10.5713/ajas.19.0889
- Fan GJ, Chen MH, Lee CF, et al. Effects of rice straw fermented with spent Pleurotus sajor-caju mushroom substrates on milking performance in Alpine dairy goats. Anim Biosci. 2022;35(7):999–1009. doi: 10.5713/ab.21.0340
- Yuan C, Wu M, Tahir SM, et al. Velvet antler production and hematological changes in male Sika deers fed with spent mushroom substrate. Animals (Basel). 2022;12(13):1689. doi: 10.3390/ani12131689
- Huang LQ, Li SQ, Yuan ZZ, et al. Effects of feeding co-fermented whole plant rice and spent mushroom(Pleurotus ostreatus) substrate on slaughter performance, meat quality and organ size indexes of Liuyang black goats. Acta Prataculturae Sin. 2021;30(6):133.
- Qi Q, Peng Q, Tang M, et al. Microbiome analysis investigating the impacts of fermented spent mushroom substrates on the composition of microbiota in weaned piglets hindgut. Front Vet Sci. 2020;7:584243. doi: 10.3389/fvets.2020.584243.
- Liu Y, Mei L, Li J, et al. Effect of JUNCAO-cultivated Ganoderma lucidum spent mushroom substrate-hot water extract on immune function in mice. Trop J Pharm Res. 2018;17(2):261–267. doi: 10.4314/tjpr.v17i2.10
- Baek YC, Kim MS, Reddy KE, et al. Rumen fermentation and digestibility of spent mushroom (Pleurotus ostreatus) substrate inoculated with Lactobacillus brevis for Hanwoo steers ¤. Revista Colombiana de Ciencias Pecuarias. 2017;30(4):267–277. doi: 10.17533/udea.rccp.v30n4a02
- Chang S, Lin MJ, Chao YP, et al. Effects of spent mushroom compost meal on growth performance and meat characteristics of grower geese. R Bras Zootec. 2016;45(6):281–287. doi: 10.1590/S1806-92902016000600001
- Seok JS, Kim YI, Lee YH, et al. Effect of feeding a by-product feed-based silage on nutrients intake, apparent digestibility, and nitrogen balance in sheep. J Anim Sci Technol. 2016;58(1):9. doi: 10.1186/s40781-016-0091-7
- Aldoori ZT, Al-Obaidi ASA, Abdulkareem AH, et al. Effect of dietary replacement of barley with mushroom cultivation on carcass characteristics of Awassi lambs. J Anim Health Prod. 2015;3(4):94–98. doi: 10.14737/journal.jahp/2015/3.4.94.98
- Kim YI, Park JM, Lee YH, et al. Effect of by-product feed-based silage feeding on the performance, blood metabolites, and carcass characteristics of Hanwoo steers (a field study). Asian-Australas J Anim Sci. 2015;28(2):180–187. doi: 10.5713/ajas.14.0443
- Liu Y, Zhao C, Lin D, et al. Effects of Ganoderma lucidum spent mushroom substrate extract on milk and serum immunoglobulin levels and serum antioxidant capacity of dairy cows. Trop J Pharm Res. 2015;14(6):1049–1055. doi: 10.4314/tjpr.v14i6.16
- Liu Y, Zhao C, Lin D, et al. Effect of water extract from spent mushroom substrate after Ganoderma balabacense cultivation by using JUNCAO technique on production performance and hematology parameters of dairy cows. Anim Sci J. 2015;86(9):855–862. doi: 10.1111/asj.12371
- Fazaeli H, Shafyee-Varzeneh H, Farahpoor A, et al. Recycling of mushroom compost wheat straw in the diet of feedlot calves with two physical forms. Int J Recycl Org Waste Agricult. 2014;3(3):3. doi: 10.1007/s40093-014-0065-z
- Foluke A, Olutayo A, Olufemi A. Assessing spent mushroom substrate as a replacement to wheat bran in the diet of broilers. Am Int J Contemp Res. 2014;4(4):178–183.
- Kim SC, Moon YH, Kim HS, et al. Effect of dietary fermented spent mushroom (Hypsizygus marmoreus) substrates on laying hens. J Mushroom. 2014;12(4):350–356. doi: 10.14480/JM.2014.12.4.350
- Tasaki Y, Kozuka K, Mochida K, et al. Effect of sawdust-based spent mushroom substrate treated with steam on rat growth performance. Food Sci Technol Res. 2014;20(2):493–497. doi: 10.3136/fstr.20.493
- Kim YI, Lee YH, Kim KH, et al. Effects of supplementing microbially-fermented spent mushroom substrates on growth performance and carcass characteristics of Hanwoo steers (a field study). Asian-Australas J Anim Sci. 2012;25(11):1575–1581. doi: 10.5713/ajas.2012.12251
- Kim MK, Lee HG, Park JA, et al. Recycling of fermented sawdust-based oyster mushroom spent substrate as a feed supplement for postweaning calves. Asian-Australas J Anim Sci. 2011;24(4):493–499. doi: 10.5713/ajas.2011.10333
- Cho YJ, Choi EJ, Shin PG, et al. Effect of spent mushroom (Pleurotus eryngii) substrates addition on egg quality in laying hens. J Mushrooms. 2010;8(4):196–197.
- Azevedo RS, Ávila CDS, Dias ES, et al. Utilization of the spent substrate of Pleurotus sajor caju mushroom in broiler chicks ration and the effect on broiler chicken performance. Acta Sci - Anim Sci. 2009;31(2):139–144.
- Machado AMB, Dias ES, dos Santos ÉC, et al. Composto exaurido do cogumelo Agaricus blazei na dieta de frangos de corte. R Bras Zootec. 2007;36(4 suppl):1113–1118. doi: 10.1590/S1516-35982007000500018
- Song YM, Lee SD, Chowdappa R, et al. Effects of fermented oyster mushroom (Pleurotus ostreats) by-product supplementation on growth performance, blood parameters and meat quality in finishing Berkshire pigs. Animal. 2007;1(2):301–307. doi: 10.1017/S1751731107683785
- Fazaeli H, Masoodi ART. Spent wheat straw compost of Agaricus bisporus mushroom as ruminant feed. Asian-Australas J Anim Sci. 2006;19(6):845–851. doi: 10.5713/ajas.2006.845
- Beattie VE, Sneddon IA, Walker N, et al. Environmental enrichment of intensive pig housing using spent mushroom compost. Anim Sci. 2001;72(1):35–42. doi: 10.1017/S1357729800055533
- Ahmed ST, Hoon JH, Mun HS, et al. Evaluation of Lactobacillus and Bacillus-based probiotics as alternatives to antibiotics in enteric microbial challenged weaned piglets. Afr J Microbiol Res. 2014;8(1):96–104. doi: 10.5897/AJMR2013.6355
- Barba-Vidal E, Martín-Orúe SM, Castillejos L. Practical aspects of the use of probiotics in pig production: A review. Lives Sci. 2019;223:84–96. doi: 10.1016/j.livsci.2019.02.017
- Kwak AM, Lee IK, Lee SY, et al. Oxalic acid from Lentinula edodes culture filtrate: antimicrobial activity on phytopathogenic bacteria and qualitative and quantitative analyses. Mycobiology. 2016;44(4):338–342. doi: 10.5941/MYCO.2016.44.4.338
- Arriola KG, Queiroz OCM, Romero JJ, et al. Effect of microbial inoculants on the quality and aerobic stability of bermudagrass round-bale haylage. J Dairy Sci. 2015;98(1):478–485. doi: 10.3168/jds.2014-8411
- Zakaria Z, Rasib NAA, Tompang MF. Spent mushroom substrate based fish feed affects the growth of catfish (Clarias gariepinus). IOP Conf Ser Earth Environ Sci. 2021;765(1):012082. doi: 10.1088/1755-1315/765/1/012082
- Van Doan H, Hoseinifar SH, Dawood MAO, et al. Effects of Cordyceps militaris spent mushroom substrate and Lactobacillus plantarum on mucosal, serum immunology and growth performance of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017;70:87–94. doi: 10.1016/j.fsi.2017.09.002
- Lee PT, Wu YS, Tseng CC, et al. Dietary Agaricus blazei spent substrate improves disease resistance of Nile tilapia (Oreochromis niloticus) against Streptococcus agalactiae in vivo. J Mar Sci Eng. 2022;10(1):100. doi: 10.3390/jmse10010100
- Chirapongsatonkul N, Mueangkan N, Wattitum S, et al. Comparative evaluation of the immune responses and disease resistance of Nile tilapia (Oreochromis niloticus) induced by yeast β-glucan and crude glucan derived from mycelium in the spent mushroom substrate of Schizophyllum commune. Aquacult Rep. 2019;15:100205. doi: 10.1016/j.aqrep.2019.100205
- Abro Z, Kassie M, Tanga C, et al. Socio-economic and environmental implications of replacing conventional poultry feed with insect-based feed in Kenya. J Clean Prod. 2020;265:121871. doi: 10.1016/j.jclepro.2020.121871
- Hawkey KJ, Lopez-Viso C, Brameld JM, et al. Insects: a potential source of protein and other nutrients for feed and food. Ann Rev Anim Biosci. 2021;9(1):333–354. doi: 10.1146/annurev-animal-021419-083930
- Roffeis M, Wakefield ME, Almeida J, et al. Life cycle cost assessment of insect based feed production in West Africa. J Clean Prod. 2018;199:792–806. doi: 10.1016/j.jclepro.2018.07.179
- Li TH, Zhang CR, Che PF, et al. Recycling of spent mushroom substrate and food waste: utilisation as feed materials for black soldier fly (Hermetia illucens (L.) Diptera: Stratiomyidae). J Ins Food Feed. 2021;7(4):409–417. doi: 10.3920/JIFF2020.0105
- Li TH, Che PF, Zhang CR, et al. Recycling of spent mushroom substrate: Utilization as feed material for the larvae of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). PLoS One. 2020 Aug 6;15(8):e0237259. doi: 10.1371/journal.pone.0237259
- Wei P, Li Y, Lai D, et al. Protaetia brevitarsis larvae can feed on and convert spent mushroom substrate from Auricularia auricula and Lentinula edodes cultivation. Waste Manag. 2020;114:234–239. doi: 10.1016/j.wasman.2020.07.009
- FAO. World fertilizer trends and outlook to 2020. Rome; 2017.
- Sharma A, Shukla A, Attri K, et al. Global trends in pesticides: A looming threat and viable alternatives. Ecotoxicol Environ Saf. 2020;201:110812. doi: 10.1016/j.ecoenv.2020.110812
- YongShan C, Camps-Arbestain M, QinHua S, et al. The long-term role of organic amendments in building soil nutrient fertility: a meta-analysis and review. Nutr Cycl Agroecosyst. 2018;111(2–3):103–125. doi: 10.1007/s10705-017-9903-5
- Celestina C, Hunt JR, Sale PWG, et al. Attribution of crop yield responses to application of organic amendments: A critical review. Soil Till Res. 2019;186:135–145. doi: 10.1016/j.still.2018.10.002
- Urra J, Alkorta I, Garbisu C. Potential benefits and risks for soil health derived from the use of organic amendments in agriculture. Agronomy. 2019;9(9):542. doi: 10.3390/agronomy9090542
- Ribas LCC, de Mendonça MM, Camelini CM, et al. Use of spent mushroom substrates from Agaricus subrufescens (syn. A. blazei, A. brasiliensis) and Lentinula edodes productions in the enrichment of a soil-based potting media for lettuce (Lactuca sativa) cultivation: Growth promotion and soil bioremediation. Bioresour Technol. 2009;100(20):4750–4757.
- Paredes C, Medina E, Bustamante MA, et al. Effects of spent mushroom substrates and inorganic fertilizer on the characteristics of a calcareous clayey-loam soil and lettuce production. Soil Use And Manag. 2016;32(4):487–494. doi: 10.1111/sum.12304
- Upadhyay SK, Chauhan PK. Optimization of eco-friendly amendments as sustainable asset for salt-tolerant plant growth-promoting bacteria mediated maize (Zea mays L.) plant growth, Na uptake reduction and saline soil restoration. Environ Res. 2022;211:113081. doi: 10.1016/j.envres.2022.113081
- Herrero-Hernández E, Andrades MS, Villalba Eguren G, et al. Organic amendment for the recovery of vineyard soils: effects of a single application on soil properties over two years. Processes. 2022;10(2):317. doi: 10.3390/pr10020317
- Hernández D, Ros M, Carmona F, et al. Composting spent mushroom substrate from Agaricus bisporus and Pleurotus ostreatus production as a growing media component for baby leaf lettuce cultivation under Pythium irregulare biotic stress. Horticulturae. 2021;7(2):13. doi: 10.3390/horticulturae7020013
- Lipiec J, Usowicz B, Kłopotek J, et al. Effects of application of recycled chicken manure and spent mushroom substrate on organic matter, acidity, and hydraulic properties of sandy soils. Materials. 2021;14(14):4036. doi: 10.3390/ma14144036
- Ngan NM, Riddech N. Use of spent mushroom substrate as an inoculant carrier and an organic fertilizer and their impacts on Roselle growth (Hibiscus sabdariffa L.) and soil quality. Waste Biomass Valor. 2021;12(7):3801–3811. doi: 10.1007/s12649-020-01278-w
- Wang HW, Xu M, Cai XY, et al. Evaluation of soil microbial communities and enzyme activities in cucumber continuous cropping soil treated with spent mushroom (Flammulina velutipes) substrate. J Soils Sediments. 2021;21(8):2938–2951. doi: 10.1007/s11368-021-02989-w
- Paula FS, Tatti E, Thorn C, et al. Soil prokaryotic community resilience, fungal colonisation and increased cross-domain co-occurrence in response to a plant-growth enhancing organic amendment. Soil Biol Biochem. 2020;149:107937. doi: 10.1016/j.soilbio.2020.107937
- Meng X, Liu B, Zhang H, et al. Co-composting of the biogas residues and spent mushroom substrate: Physicochemical properties and maturity assessment. Bioresour Technol. 2019;276:281–287. doi: 10.1016/j.biortech.2018.12.097
- Ultra VU, Sotto JO, Punzalan MR. Nutrient supplying potential of different spent mushroom substrate preparations as soil amendment in a potting media. IOP Conf Ser Earth Environ Sci. 2018;120:012010. doi: 10.1088/1755-1315/120/1/012010
- Paula FS, Tatti E, Abram F, et al. Stabilisation of spent mushroom substrate for application as a plant growth-promoting organic amendment. J Environ Manage. 2017;196:476–486. doi: 10.1016/j.jenvman.2017.03.038
- Gobbi V, Bonato S, Nicoletto C, et al. Spent mushroom substrate as organic fertilizer: vegetable organic trials | International society for horticultural science. Proceedings of the III International Symposium on Organic Matter Management and Compost Use in Horticulture. Murcia, Spain; 2015. p. 49–56.
- Nakatsuka H, Oda M, Hayashi Y, et al. Effects of fresh spent mushroom substrate of Pleurotus ostreatus on soil micromorphology in Brazil. Geoderma. 2016;269:54–60. doi: 10.1016/j.geoderma.2016.01.023
- Sendi H, Mohamed MTM, Anwar MP, et al. Spent mushroom waste as a media replacement for peat moss in Kai-Lan (Brassica oleracea var. alboglabra) production. Sci World J. 2013;2013:1–8. doi: 10.1155/2013/258562
- Medina E, Paredes C, Bustamante MA, et al. Relationships between soil physico-chemical, chemical and biological properties in a soil amended with spent mushroom substrate. Geoderma. 2012;173–174:152–161. doi: 10.1016/j.geoderma.2011.12.011
- Peregrina F, Larrieta C, Colina M, et al. Spent mushroom substrates influence soil quality and nitrogen availability in a semiarid vineyard soil. Soil Sci Soc Am J. 2012;76(5):1655–1666. doi: 10.2136/sssaj2012.0018
- Wuest PJ, Fahy HK, Fahy J. Use of spent mushroom substrate (SMS) for corn (maize) production and its effect on surface water quality. Compost Sci Util. 1995;3(1):46–54. doi: 10.1080/1065657X.1995.10701768
- Meng L, Li W, Zhang S, et al. Feasibility of co-composting of sewage sludge, spent mushroom substrate and wheat straw. Bioresour Technol. 2017;226:39–45. doi: 10.1016/j.biortech.2016.11.054
- Meng L, Zhang S, Gong H, et al. Improving sewage sludge composting by addition of spent mushroom substrate and sucrose. Bioresour Technol. 2018;253:197–203. doi: 10.1016/j.biortech.2018.01.015
- Meng X, Liu B, Xi C, et al. Effect of pig manure on the chemical composition and microbial diversity during co-composting with spent mushroom substrate and rice husks. Bioresour Technol. 2018;251:22–30. doi: 10.1016/j.biortech.2017.09.077
- Kong Y, Ma R, Li G, et al. Impact of biochar, calcium magnesium phosphate fertilizer and spent mushroom substrate on humification and heavy metal passivation during composting. Sci Total Environ. 2022;824:153755. doi: 10.1016/j.scitotenv.2022.153755
- Liu Y, Ma R, Li D, et al. Effects of calcium magnesium phosphate fertilizer, biochar and spent mushroom substrate on compost maturity and gaseous emissions during pig manure composting. Journal Of Environmental Management query. 2020;267:110649–. doi: 10.1016/j.jenvman.2020.110649
- Yu X, Li X, Ren C, et al. Co-composting with cow dung and subsequent vermicomposting improve compost quality of spent mushroom. Bioresour Technol. 2022;358:127386. doi: 10.1016/j.biortech.2022.127386
- Sun C, Wei Y, Kou J, et al. Improve spent mushroom substrate decomposition, bacterial community and mature compost quality by adding cellulase during composting. J Clean Prod. 2021;299:126928. doi: 10.1016/j.jclepro.2021.126928
- Domínguez-Gutiérrez M, Gaitán-Hernández R, Moctezuma-Pérez I, et al. Composting and vermicomposting of spent mushroom substrate to produce organic fertilizer. Emir J Food Agric. 2022;34(3):220–228. doi: 10.9755/ejfa.2022.v34.i3.2828
- Stack JP, Kenerley CM, Pettit RE. Application of biological control agents. In: Biocontrol of plant diseases. CRC Press; 2020. pp. 43–54.
- Wright MG, Bennett GM. Evolution of biological control agents following introduction to new environments. BioControl. 2018;63(1):105–116. doi: 10.1007/s10526-017-9830-z
- Shen HS, Shao S, Chen JC, et al. Antimicrobials from mushrooms for assuring food safety. Compr Rev Food Sci Food Saf. 2017;16(2):316–329. doi: 10.1111/1541-4337.12255
- Katas H, Akhmar MAM, Abdalla SSI. Biosynthesized silver nanoparticles loaded in gelatine hydrogel for a natural antibacterial and anti-biofilm wound dressing. J Bioact Compat Polym. 2021;36(2):111–123. doi: 10.1177/0883911520988303
- Velazhahan R, Al-Mamari SNH, Al-Sadi AM, et al. In vitro antagonistic potential, plant growth-promoting activity and indole-3-acetic acid producing trait of bacterial isolates from spent mushroom substrate of Agaricus bisporus. Jour Agri Mar Scie. 2020;25(2):22–29. doi: 10.24200/jams.vol25iss2pp22-29
- Li H, Yoshida S, Mitani N, et al. Disease resistance and growth promotion activities of chitin/cellulose nanofiber from spent mushroom substrate to plant. Carbohydr Polym. 2022;284:119233. doi: 10.1016/j.carbpol.2022.119233
- Fujita R, Yokono M, Ube N, et al. Suppression of Alternaria brassicicola infection by volatile compounds from spent mushroom substrates. J Biosci Bioeng. 2021;132(1):25–32. doi: 10.1016/j.jbiosc.2021.03.003
- Singh G, Tiwari A, Gupta A, et al. Bioformulation development via valorizing silica-rich spent mushroom substrate with Trichoderma asperellum for plant nutrient and disease management. J Environ Manage. 2021;297:113278. doi: 10.1016/j.jenvman.2021.113278
- Yu YY, Li SM, Qiu JP, et al. Combination of agricultural waste compost and biofertilizer improves yield and enhances the sustainability of a pepper field. J Plant Nutr Soil Sci. 2019;182(4):560–569. doi: 10.1002/jpln.201800223
- Aguiar T, Luiz C, Neto A, et al. Residual polysaccharides from fungi reduce the bacterial spot in tomato plants. Bragantia. 2018;77(2):299–313. doi: 10.1590/1678-4499.2016514
- Biswas MK, Ghosh T. Screening of brinjal genotypes for their resistance against fungal and bacterial wilt and integrated management of the disease. Plant Cell Biotechnol Mol Biol. 2018;19:61–71.
- Ishihara A, Goto N, Kikkawa M, et al. Identification of antifungal compounds in the spent mushroom substrate of Lentinula edodes. J Pestic Sci. 2018;43(2):108–113.
- Kang DS, Min KJ, Kwak AM, et al. Defense response and suppression of phytophthora blight disease of pepper by water extract from spent mushroom substrate of Lentinula edodes. Plant Pathol J. 2017;33(3):264–275. doi: 10.5423/PPJ.OA.02.2017.0030
- Chen JT, Lin MJ, Huang JW. Efficacy of spent blewit mushroom compost and Bacillus aryabhattai combination on control of Pythium damping-off in cucumber. J Agric Sci. 2015;153(7):1257–1266. doi: 10.1017/S0021859614000987
- Kwak AM, Min KJ, Lee SY, et al. Water extract from spent mushroom substrate of Hericium erinaceus suppresses bacterial wilt disease of tomato. Mycobiology. 2015;43(3):311–318. doi: 10.5941/MYCO.2015.43.3.311
- Nicol RW, Burlakoti P. Effect of aerobic compost tea inputs and application methods on protecting tomato from Phytophthora capsici. Acta Hortic. 2015;1069(1069):229–233. doi: 10.17660/ActaHortic.2015.1069.32
- Marín F, Santos M, Diánez F, et al. Characters of compost teas from different sources and their suppressive effect on fungal phytopathogens. World J Microbiol Biotechnol. 2013;29(8):1371–1382. doi: 10.1007/s11274-013-1300-x
- Parada RY, Murakami S, Shimomura N, et al. Suppression of fungal and bacterial diseases of cucumber plants by using the spent mushroom substrate of Lyophyllum decastes and Pleurotus eryngii. J Phytopathol. 2012;160(7–8):390–396. doi: 10.1111/j.1439-0434.2012.01916.x
- Mansour F, El-Sayed G. Soil amendment and seed treatments with compost tea as alternative fungicide for controlling root rot disease of bean plants - proquest. Egypt J Biol Pest Control. 2011;21(1):19–26.
- Parada RY, Murakami S, Shimomura N, et al. Autoclaved spent substrate of hatakeshimeji mushroom (Lyophyllum decastes Sing.) and its water extract protect cucumber from anthracnose. Crop Prot. 2011;30(4):443–450. doi: 10.1016/j.cropro.2010.11.021
- Ntougias S, Papadopoulou KK, Zervakis GI, et al. Suppression of soil-borne pathogens of tomato by composts derived from agro-industrial wastes abundant in mediterranean regions. Biol Fertil Soils. 2008;44(8):1081–1090. doi: 10.1007/s00374-008-0295-1
- Kaur H, Nyochembeng LM, Mentreddy SR, et al. Assessment of the antimicrobial activity of Lentinula edodes against Xanthomonas campestris pv. Vesicatoria. Crop Prot. 2016;89:284–288. doi: 10.1016/j.cropro.2016.08.001
- Elsakhawy T, ALKahtani MDF, Sharshar AAH, et al. Efficacy of mushroom metabolites (Pleurotus ostreatus) as a natural product for the suppression of broomrape growth (Orobanche crenata Forsk) in faba bean plants. Plants. 2020;9(10):1265. doi: 10.3390/plants9101265
- Elsakhawy T, Abd El-Rahem W. Evaluation of spent mushroom substrate extract as a biofertilizer for growth improvement of rice (Oryza sativa L.). Egypt J Soil Sci. 2020;60(1):31–42.
- Gibbs J, Gaskin E, Ji C, et al. The effect of plant-based dietary patterns on blood pressure: a systematic review and meta-analysis of controlled intervention trials. J Hypertens. 2021;39(1):23–37. doi: 10.1097/HJH.0000000000002604
- Yu E, Malik VS, Hu FB. Cardiovascular disease prevention by diet modification: JACC health promotion series. J Am Coll Cardiol. 2018;72(8):914–926. doi: 10.1016/j.jacc.2018.02.085
- Thakur M, Bhattacharya S, Khosla PK, et al. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J Appl Res Med Aromat Plants. 2019;12:1–12. doi: 10.1016/j.jarmap.2018.11.004
- Vahid Afagh H, Saadatmand S, Riahi H, et al. Influence of spent mushroom compost (SMC) as an organic fertilizer on nutrient, growth, yield, and essential oil composition of German chamomile (Matricaria recutita L.). Commun Soil Sci Plant Anal. 2019;50(5):538–548. doi: 10.1080/00103624.2019.1568450
- Vahid Afagh H, Saadatmand S, Riahi H, et al. Effects of Leached Spent Mushroom Compost (LSMC) on the yield, essential oil composition and antioxidant compounds of German chamomile (Matricaria recutita L.). J Essent Oil Bear Plants. 2018;21(6):1436–1449. doi: 10.1080/0972060X.2019.1580617
- Esmaielpour B, Rahmanian M, Heidarpour O, et al. Effect of vermicompost and spent mushroom compost on the nutrient and essential oil composition of Basil (Ocimum basilicum L.). J Essent Oil Bear Plants. 2017;20(5):1283–1292. doi: 10.1080/0972060X.2017.1396931
- Roy S, Barman S, Chakraborty UCB. Evaluation of spent mushroom substrate as biofertilizer for growth improvement of Capsicum annuum L. J App Biol Biotech. 2015;3(3):022–7.
- Singh UB, Malviya D, Khan W, et al. Earthworm grazed-Trichoderma harzianum biofortified spent mushroom substrates modulate accumulation of natural antioxidants and bio-fortification of mineral nutrients in tomato. Front Plant Sci. 2018;9:1017. doi: 10.3389/fpls.2018.01017
- Medina E, Paredes C, Pérez-Murcia MD, et al. Spent mushroom substrates as component of growing media for germination and growth of horticultural plants. Bioresour Technol. 2009;100(18):4227–4232. doi: 10.1016/j.biortech.2009.03.055
- Ariff INM, Bahrin EK, Ramli N, et al. Direct use of spent mushroom substrate from Pleurotus pulmonarius as a readily delignified feedstock for cellulase production. Waste Biomass Valori. 2019;10(4):839–850. doi: 10.1007/s12649-017-0106-8
- Schallemberger JB, Libardi N, Dalari BLSK, et al. Textile azo dyes discolouration using spent mushroom substrate: enzymatic degradation and adsorption mechanisms. Environmental Technology. 2021;44(9):1265–1286. doi: 10.1080/09593330.2021.2000038
- Lau KL, Tsang YY, Chiu SW. Use of spent mushroom compost to bioremediate PAH-contaminated samples. Chemosphere. 2003;52(9):1539–1546. doi: 10.1016/S0045-6535(03)00493-4
- Ball AS, Jackson AM. The recovery of lignocellulose-degrading enzymes from spent mushroom compost. Bioresour Technol. 1995;54(3):311–314. doi: 10.1016/0960-8524(95)00153-0
- Trejo-Hernandez MR, Lopez-Munguia A, Ramirez RQ. Residual compost of Agaricus bisporus as a source of crude laccase for enzymic oxidation of phenolic compounds. Process Biochem. 2001;36(7):635–639. doi: 10.1016/S0032-9592(00)00257-0
- Mayolo-Deloisa K, Del Refugio Trejo-Hernandez M, Rito-Palomares M. Recovery of laccase from the residual compost of Agaricus bisporus in aqueous two-phase systems. Process Biochemistry query. 2009;44(4):435–439. doi: 10.1016/j.procbio.2008.12.010
- Devi R, Kapoor S, Thakur R, et al. Lignocellulolytic enzymes and bioethanol production from spent biomass of edible mushrooms using Saccharomyces cerevisiae and Pachysolen tannophilus. Biomass Convers Biorefin. 2022;1–15. doi: 10.1007/s13399-022-02406-3
- Singh AD, Abdullah N, Vikineswary S. Optimization of extraction of bulk enzymes from spent mushroom compost. J Chem Technol Biotechnol. 2003;78(7):743–752. doi: 10.1002/jctb.852
- Ko HG, Park SH, Kim SH, et al. Detection and recovery of hydrolytic enzymes from spent compost of four mushroom species. Folia Microbiol. 2005;50(2):103. doi: 10.1007/BF02931456
- Lim SH, Lee YH, Kang HW. Efficient recovery of lignocellulolytic enzymes of spent mushroom compost from oyster mushrooms, Pleurotus spp., and potential use in dye decolorization. Mycobiology. 2013;41(4):214–220. doi: 10.5941/MYCO.2013.41.4.214
- Lim SH, Kim JK, Lee YH. Production of lignocellulolytic enzymes from spent mushroom compost of Pleurotus eryngii. Korean J Mycol. 2012;40(3):152–158. doi: 10.4489/KJM.2012.40.3.152
- Sadiq S, Hassan MM, Saba S, et al. Bioremediation of endosulfan in soil using ligninolytic extract of spent mushroom compost of Pleurotus ostreatus. Appl Ecol Environ Res. 2019;17(2):3251–3267. doi: 10.15666/aeer/1702_32513267
- Rajavat AS, Rai S, Pandiyan K, et al. Sustainable use of the spent mushroom substrate of Pleurotus florida for production of lignocellulolytic enzymes. J Of Basic Microbiol. 2020;60(2):173–184. doi: 10.1002/jobm.201900382
- Yunan NAM, Shin TY, Ibrahim S, et al. Upcycling the spent mushroom substrate of the grey oyster mushroom Pleurotus pulmonarius as a source of lignocellulolytic enzymes for palm oil mill effluent hydrolysis. J Microbiol Biotechnol. 2021;31(6):823–832. doi: 10.4014/jmb.2103.03020
- Lin H, Sun M, Li J, et al. Purification and characterization of xylanase from spent mushroom compost and its application in saccharification of biomass wastes. BioResources. 2017;13(1):220–230. doi: 10.15376/biores.13.1.220-230
- Schimpf U, Schulz R. Industrial by-products from white-rot fungi production. Part I: Generation of enzyme preparations and chemical, protein biochemical and molecular biological characterization. Process Biochem. 2016;51(12):2034–2046. doi: 10.1016/j.procbio.2016.08.032
- Nakajima VM, Soares FEF, Queiroz JH. Screening and decolorizing potential of enzymes from spent mushroom composts of six different mushrooms. Biocatal Agric Biotechnol. 2018;13:58–61. doi: 10.1016/j.bcab.2017.11.011
- Liao CS, Yuan SY, Hung BH, et al. Removal of organic toxic chemicals using the spent mushroom compost of Ganoderma lucidum. J Environ Monit. 2012;14(7):1983–1988. doi: 10.1039/c2em10910g
- Phan CW, Sabaratnam V. Potential uses of spent mushroom substrate and its associated lignocellulosic enzymes. Appl Microbiol Biotechnol. 2012;96(4):863–873. doi: 10.1007/s00253-012-4446-9
- Branà MT, Sergio L, Haidukowski M, et al. Degradation of aflatoxin B1 by a sustainable enzymatic extract from spent mushroom substrate of Pleurotus eryngii. Toxins (Basel). 2020;12(1):49. doi: 10.3390/toxins12010049
- Passos DF, Pereira N, Castro AM. A comparative review of recent advances in cellulases production by Aspergillus, Penicillium and Trichoderma strains and their use for lignocellulose deconstruction. Curr Opin Green Sustain Chem. 2018;14:60–66. doi: 10.1016/j.cogsc.2018.06.003
- Feng Y, Liu HQ, Xu F, et al. Enzymatic degradation of steam-pretreated Lespedeza stalk (Lespedeza crytobotrya) by cellulosic-substrate induced cellulases. Bioproc Biosyst Eng. 2011;34:357–365. doi: 10.1007/s00449-010-0478-7
- Martín C, Dixit P, Momayez F, et al. Hydrothermal pretreatment of lignocellulosic feedstocks to facilitate biochemical conversion. Front Bioeng Biotechnol. 2022;10:10. doi: 10.3389/fbioe.2022.846592
- Jönsson LJ, Martín C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour Technol. 2016;199:103–112. doi: 10.1016/j.biortech.2015.10.009
- Chen F, Xiong S, Gandla ML, et al. Spent mushroom substrates for ethanol production – effect of chemical and structural factors on enzymatic saccharification and ethanolic fermentation of Lentinula edodes-pretreated hardwood. Bioresour Technol. 2022;347:126381. doi: 10.1016/j.biortech.2021.126381
- He J, Qiu Y, Ji X, et al. A novel strategy for producing cellulase from Trichoderma reesei with ultrasound‐assisted fermentation using spent mushroom substrate. Process Biochem. 2021;104:110–116. doi: 10.1016/j.procbio.2021.03.015
- Grujić M, Dojnov B, Potočnik I, et al. Spent mushroom compost as substrate for the production of industrially important hydrolytic enzymes by fungi Trichoderma spp. and Aspergillus niger in solid state fermentation. Int Biodeter Biodegr. 2015;104:290–298. doi: 10.1016/j.ibiod.2015.04.029
- Camassola M, Dillon AJP. Biological pretreatment of sugar cane bagasse for the production of cellulases and xylanases by Penicillium echinulatum. Ind Crops Prod. 2009;29(2):642–647. doi: 10.1016/j.indcrop.2008.09.008
- Economou CN, Diamantopoulou PA, Philippoussis AN. Valorization of spent oyster mushroom substrate and laccase recovery through successive solid state cultivation of Pleurotus, Ganoderma, and Lentinula strains. Appl Microbiol Biotechnol. 2017;101(12):5213–5222. doi: 10.1007/s00253-017-8251-3
- Kumar K, Mehra R, Guiné RPF, et al. Edible mushrooms: a comprehensive review on bioactive compounds with health benefits and processing aspects. Foods. 2021;10(12):2996. doi: 10.3390/foods10122996
- Zhang Y, Liu W, Huang W, et al. Characterization and antiproliferative effect of novel acid polysaccharides from the spent substrate of shiitake culinary-medicinal mushroom Lentinus edodes (Agaricomycetes) cultivation. Int J Medic Mushrooms. 2017;19(5):395–403. doi: 10.1615/IntJMedMushrooms.v19.i5.20
- Liu Y, Zhen D, Wang D, et al. Immunomodulatory activities of polysaccharides from white button mushroom, Agaricus bisporus (Agaricomycetes), fruiting bodies and cultured mycelia in healthy and immunosuppressed mice. Int J Medic Mushrooms. 2019;21(1):13–27. doi: 10.1615/IntJMedMushrooms.2018029648
- Zhu H, Sheng K, Yan E, et al. Extraction, purification and antibacterial activities of a polysaccharide from spent mushroom substrate. Int j biol macromol. 2012;50(3):840–843. doi: 10.1016/j.ijbiomac.2011.11.016
- Ren Z, Liu W, Song X, et al. Antioxidant and anti-inflammation of enzymatic-hydrolysis residue polysaccharides by Lentinula edodes. Int j biol macromol. 2018;120:811–822. doi: 10.1016/j.ijbiomac.2018.08.114
- Zhang J, Meng G, Zhai G, et al. Extraction, characterization and antioxidant activity of polysaccharides of spent mushroom compost of Ganoderma lucidum. Int j biol macromol. 2016;82:432–439. doi: 10.1016/j.ijbiomac.2015.10.016
- Dávila-Giraldo LR, Zambrano-Forero C, Torres-Arango O, et al. Integral use of rice husks for bioconversion with white-rot fungi. Biomass Convers Biorefin. 2022;12(8):2981–2991. doi: 10.1007/s13399-020-00940-6
- He P, Li F, Huang L, et al. Chemical characterization and antioxidant activity of polysaccharide extract from spent mushroom substrate of Pleurotus eryngii. J Taiwan Inst Chem Eng. 2016;69:48–53. doi: 10.1016/j.jtice.2016.10.017
- Zeng Y, Zhang Y, Zhang L, et al. Structural characterization and antioxidant and immunomodulation activities of polysaccharides from the spent rice substrate of Cordyceps militaris. Food Sci Biotechnol. 2015;24(5):1591–1596. doi: 10.1007/s10068-015-0205-x
- Zhang Y, Zeng Y, Zhang LJ, et al. Monosaccharide composition and antioxidant activity of polysaccharides from different spent mushroom substrates. Food Sci. 2015;36(5):18–23.
- Zou G, Li B, Wang Y, et al. Efficient conversion of spent mushroom substrate into a high value-added anticancer drug pentostatin with engineered Cordyceps militaris. Green Chem. 2021;23(24):10030–10038. doi: 10.1039/D1GC03594K
- Umaña M, Eim V, Garau C, et al. Ultrasound-assisted extraction of ergosterol and antioxidant components from mushroom by-products and the attainment of a β-glucan rich residue. Food Chem. 2020;332:127390. doi: 10.1016/j.foodchem.2020.127390
- Vis M, Mantau U, Allen B, Eds. Study on the optimised cascading use of wood Figure 3. https://data.europa.eu/doi/10.2873/827106
- Keegan D, Kretschmer B, Elbersen B, et al. Cascading use: a systematic approach to biomass beyond the energy sector. Biofuels Bioprod Biorefin. 2013;7(2):193–206. doi: 10.1002/bbb.1351
- Chen F, Martín C, Lestander TA, et al. Shiitake cultivation as biological preprocessing of lignocellulosic feedstocks – substrate changes in crystallinity, syringyl/guaiacyl lignin and degradation-derived by-products. Bioresour Technol. 2022;344:126256. doi: 10.1016/j.biortech.2021.126256
- Xiong S, Martín C, Eilertsen L, et al. Energy-efficient substrate pasteurisation for combined production of shiitake mushroom (Lentinula edodes) and bioethanol. Bioresour Technol. 2019;274:65–72. doi: 10.1016/j.biortech.2018.11.071
- Chen F Combined production of edible mushrooms and biofuels from lignocellulosic residues [Internet] [ Doctoral thesis]. 2021 [cited 2022 Aug 22]. Available from: https://pub.epsilon.slu.se/26324/1/chen_f_211216.pdf
- Chen F, Grimm A, Eilertsen L, et al. Integrated production of edible mushroom (Auricularia auricula-judae), fermentable sugar and solid biofuel. Renewable Energy. 2021;170:172–180. doi: 10.1016/j.renene.2021.01.124
- Lin Y, Ge X, Liu Z, et al. Integration of shiitake cultivation and solid-state anaerobic digestion for utilization of woody biomass. Bioresour Technol. 2015;182:128–135. doi: 10.1016/j.biortech.2015.01.102
- Schimpf U, Schulz R. Industrial by-products from white-rot fungi production. Part II: Application in anaerobic digestion for enzymatic treatment of hay and straw. Process Biochem. 2019;76:142–154. doi: 10.1016/j.procbio.2018.10.006
- Khan MU, Ahring BK. Lignin degradation under anaerobic digestion: Influence of lignin modifications -A review. Biomass Bioenerg. 2019;128:105325. doi: 10.1016/j.biombioe.2019.105325
- Huang W, Wachemo AC, Yuan H, et al. Full utilization of nutrients in rice straw by integrating mushroom cultivation, biogas production, and fertilizer use. Int J Agric Biol Eng. 2019;12(4):174–183. doi: 10.25165/j.ijabe.20191204.4658
- Ikeda S, Yamauchi M, Watari T, et al. Development of enokitake (Flammulina velutipes) mushroom cultivation technology using spent mushroom substrate anaerobic digestion residue. Environ Technol Innov. 2021;24:102046. doi: 10.1016/j.eti.2021.102046
- Elisashvili V, Penninckx M, Kachlishvili E, et al. Lentinus edodes and Pleurotus species lignocellulolytic enzymes activity in submerged and solid-state fermentation of lignocellulosic wastes of different composition. Bioresour Technol. 2008;99(3):457–462. doi: 10.1016/j.biortech.2007.01.011
- Li W, Khalid H, Zhu Z, et al. Methane production through anaerobic digestion: Participation and digestion characteristics of cellulose, hemicellulose and lignin. Appl Energy. 2018;226:1219–1228. doi: 10.1016/j.apenergy.2018.05.055
- van Kuijk SJA, Sonnenberg ASM, Baars JJP, et al. Fungal treatment of lignocellulosic biomass: Importance of fungal species, colonization and time on chemical composition and in vitro rumen degradability. Anim Feed Sci Technol. 2015;209:40–50. doi: 10.1016/j.anifeedsci.2015.07.026
- Ferreira JM, Braga FR, Soares FEF. Nematicidal activity of the Lentinula edodes’ spent mushroom compost. S Afr J Bot. 2022;146:101–102. doi: 10.1016/j.sajb.2021.10.008
- Yagüe-Carrasco MR, Lobo MC. Reutilización del sustrato post-cultivo de hongos en semillero de hortícolas. ITEA. 2021;117:347–359. doi: 10.12706/itea.2021.004
- Yu H, Liu P, Shan W, et al. Remediation potential of spent mushroom substrate on Cd pollution in a paddy soil. Environ Sci Pollut Res. 2021;28(27):36850–36860. doi: 10.1007/s11356-021-13266-1
- Chen WS, Tsai WT, Lin YQ, et al. Production of highly porous biochar materials from spent mushroom composts. Horticulturae. 2022;8(1):46. doi: 10.3390/horticulturae8010046