
ABSTRACT
Intravital imaging by multiphoton microscopy is a powerful tool to gain invaluable insight into tissue biology and function. Here, we provide a step-by-step tissue preparation protocol for imaging the mouse tibialis anterior skeletal muscle. Additionally, we include steps for jugular vein catheterization that allow for well-controlled intravenous reagent delivery. Preparation of the tibialis anterior muscle is minimally invasive, reducing the chances of inducing damage and inflammation prior to imaging. The tibialis anterior muscle is useful for imaging leukocyte interaction with vascular endothelium, and to understand muscle contraction biology. Importantly, this model can be easily adapted to study neuromuscular diseases and myopathies.
Introduction
Multiphoton microscopy is a powerful tool that allows us to conduct direct in vivo intravital imaging with high spatio-temporal resolution. This provides us with invaluable insight into cellular dynamics, tissue morphology and function that cannot be achieved easily with traditional methods like histology. Traditionally, the muscle is a very suitable imaging site for direct visualization of microcirculation due to its accessible and dense capillary bed.Citation1,2 Two commonly used muscle tissues in imaging are the cremaster muscle that covers the testicles,Citation3 and the tibialis anterior skeletal muscle.Citation4,5 Our group has previously used the tibialis anterior muscle to visualize the interaction of leukocytes with the vascular endothelium. Specifically, we demonstrated the role of CXCR4 in neutrophil margination in capillary beds with the administration of G-CSF or plerixafor.Citation6
Besides visualizing the capillary bed in the muscle tissue, other reports have also utilized tibialis anterior skeletal muscle imaging to investigate skeletal muscle tissue biology itself. The muscle tissue is rich in mitochondria to enable its contractile function,Citation1,7 and endogenous NADPH fluorescence was measured using 2-photon microscopy as a measure of mitochondrial metabolism.Citation8 A better understanding of the processes involved in mitochondria function can shed light on metabolic myopathies. Recently, multiphoton microscopy has also been adapted to image large 3D volumes of images, to reveal intricate interactions and relationships between capillaries, mitochondria and muscle fibers in sarcolemma in mice.Citation9
The cremaster muscle preparation has been widely used as a model of choice for intravital studies of leukocyte transmigration and vascular function.Citation10-12 However, the curvature of the muscle limits the observation region and exteriorization of the muscle is essential. This procedure involves detaching the muscle from the testicle and spreading it as a flat sheet for imaging. Even with the most careful exteriorization of the muscle, such surgical manipulation still results in trauma and alters the vascular permeability.Citation13,14 As a result, only a small central area distant from the manipulated circumference can be imaged to avoid artifacts that result from the surgical procedures. In contrast, the tibialis anterior skeletal muscle imaging model described here involves minimal surgical manipulation and hence, the available imaging surface is much larger and multiple regions can be examined with one single preparation.
In this report, we describe the steps required for jugular vein catheterization, and the preparation of the tibialis anterior muscle for multiphoton live imaging. Jugular vein catheterization permits fine control over the timing of intravenous reagent delivery. Hence, this permits easy visualization of early changes at the imaging site with multiphoton microscopy immediately after or during delivery of agents. To illustrate the applicability of this model, we demonstrate the use of the tibialis anterior muscle imaging to visualize how neutrophils respond to an inflammatory stimulus with lipopolysaccharide (LPS). The high spatio-temporal resolution by multiphoton microscopy combined with this muscle imaging model can be further applied to understand the biology of the outer connective tissue surrounding the skeletal muscle and muscle contraction in the context of development and disease.
Results
Insertion of jugular vein catheter for intravenous administration of drugs/labeling reagents
We anesthetize the mouse by administering ketamine/xylazine mix of 150 mg/kg ketamine and 10 mg/kg xylazine via intraperitoneal injection. Next, the heating pad is warmed up to 37°C to allow the mouse to maintain its body temperature during the preparation procedure. When the mouse is deeply anesthetized, we position the mouse with its back on the heating pad, its head pointing toward the surgeon. The four paws of the mouse are secured down on the heating pad with masking tape. We then pass a suture through the back of the top incisors of the mouse, and the suture is anchored down on the heating pad to stretch the skin of the ventral neck region ().
Figure 1. Setup for jugular vein catheterization procedure. (A) Mouse limbs and head are securely taped down to the board. (B) Small window of skin is removed at the neck region to expose the jugular vein. (C) The vein is clearly exposed (arrow) after the surrounding muscle and fat tissues are teased away. (D) Two sutures are passed underneath the jugular vein. A knot is tied with the bottom suture at the caudal end (labeled ‘cau’) to stop the blood flow. A loose knot is tied with the top suture at the rostral end (labeled ‘ros’), which will be tightened at the end of the procedure. (E) The 30G needle tip (arrow) is used to guide the insertion of the catheter (arrowhead) into the jugular vein. (F) The catheter is properly inserted into the vein and the needle tip is removed. (G) Close up of the catheter inserted into the vein. Arrow indicates the location where the catheter is inserted into the jugular vein. (H) The catheter is firmly secured with knots using both sutures. Backflow of blood in the catheter after drawing back the syringe indicates that the catheter is well inserted.
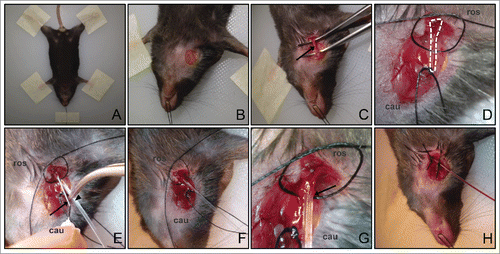
Next, we shave the hair on the ventral neck region of the mouse with a razor to expose the skin for surgery. We recommend catheterizing the right jugular vein of the mouse as the parotid gland partially obstructs the left jugular vein. We make a small incision at the skin of the ventral neck (), and gently tease away the muscle and fat tissues from the jugular vein with forceps, leaving about 5 mm of the jugular vein clearly exposed with no obstructing tissue above and underneath the vein (arrow, ).
Two strands of surgical suture (10 cm long each) are then passed underneath the jugular vein. With one thread, we tie a single overhand knot tightly near the rostral end (labeled ‘ros’ in ) of the exposed vein. The thread is then secured on the heating pad with masking tape in order to keep the vein taut, to facilitate the insertion of the catheter. With the second suture thread, we tie a loose single overhand knot near the caudal end (labeled ‘cau’ in ) of the exposed vein.
A 30G needle is bent at the halfway point at a 45° angle, and a catheter (35–40 cm long) is cut at one end to obtain a pointed end for easy insertion into the vein in the next step. We then connect the catheter to a tuberculin syringe (fitted with a 30G needle) filled with PBS, ensuring that there are no air bubbles present. Next, we place the mouse under the dissecting microscope to view the exposed jugular vein, and insert the needle tip into and parallel to the vein. When the needle tip is in the vein, lift up the needle slightly to allow the insertion of the catheter into the vein (). Remove the needle when the catheter is properly inserted (). We gently draw back the plunger of the syringe; if the catheter is positioned correctly, there will be a backflow of blood into the catheter (). Once the catheter is well positioned, we tie off the top thread tightly with a double overhand knot around the catheter to fix its position, and tie another single overhand knot for the bottom suture over the catheter to further secure the catheter. Cut off all the spare ends of the sutures (), and the syringe can then be replaced with a new one that contains the desired reagent to be introduced intravenously in the mouse.
Preparation of the tibialis anterior muscle for intravital imaging
Prepare a block of blu-tack about 2 cm × 2 cm × 0.5 cm. With the back end of forceps, make 2 indentations in the blu-tack block to rest the tibia and the foot within these grooves. Prepare two small strips of blu-tack about 2 cm long. Additionally, prepare vacuum grease filled in a 10 ml syringe for easy dispensing ().
Figure 2. Tibialis anterior skeletal muscle preparation for multiphoton intravital imaging. (A) Materials prepared: a block of blu-tack with the limb indents (dotted lines), 2 strips of blu-tack for securing the limb, and a 10 ml syringe filled with vacuum grease. (B) Right tibia and thigh are cleanly shaved with a razor. Arrow and dotted line indicate the lateral saphenous vein that runs from the ankle to the tibialis posterior muscle. (C) A small window in the skin is made to reveal the muscle tissue. Notice that the right side of the tissue from the major connective tissue (visible as a vertical white line; arrow) has a lower density of blood vessels compared to the left side. (D) Mouse leg is secured with the 2 strips of blu-tack, one placed perpendicular to the other, over the ankle. Forceps were used to press down the blu-tack strips to the block. (E) View of whole animal with the right leg placed in the mold for imaging, and the left leg taped on the heating pad. (F) A small window (dotted line) is made in the outer connective tissue to reveal the skeletal muscle tissue for imaging. The window is made to the right of the major connective tissue (visible as a vertical white line; arrow). (G) A ring of vacuum grease is drawn around the imaging area (arrow). (H) The ring of vacuum grease is filled with a drop of PBS and a coverslip mounted on the coverslip holder is lowered over the tissue. The curved holder for the thermistor (top arrowhead) and the heater block (bottom arrowhead) are screwed on to the coverslip holder. (I) Overview of the final setup for the tibialis anterior skeletal muscle imaging.
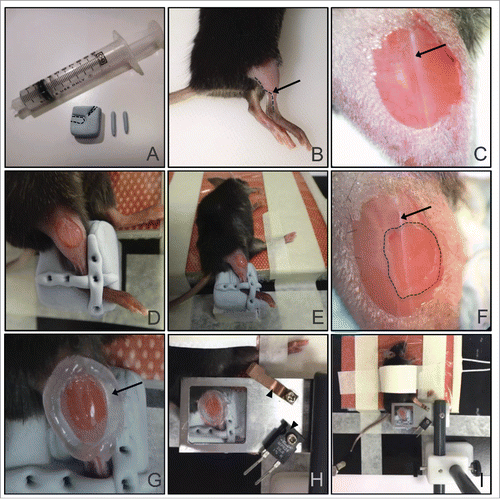
If jugular vein catheterization was performed, remove all the tapes from the heating pad, but leaving the mouse on the heating pad to continue to maintain its body temperature at 37°C during the muscle preparation. During this time, ensure that the mouse is still well anesthetized and top up with ketamine/xylazine mix if necessary. Lay the mouse in prone position.
Next, shave the right tibia and thigh cleanly with a razor (). The lateral saphenous vein, a major blood vessel that runs from the ankle to the tibialis posterior muscle, should be visible through the skin (arrow, ). We make a small incision to the right of this vein by lifting up a piece of skin with forceps and making a cut parallel to the thigh to get an exposed window of approximately 0.5 cm in length and 0.5 cm in width (). If performed carefully, this should result in minimal bleeding.
Once the incision is made, it is important that the following steps are done quickly until the coverslip is placed over the imaging area, as the exposed tissue will dry up easily. One can gently moisten the tissue area with a cotton tip applicator moistened with PBS, if necessary. We transfer the mouse on the heating pad of the imaging stage at this point, placing the leg in an extended position in the indentations of the pre-molded blu-tack (). We secure the leg in position by placing the 2 strips of blu-tack for support: the first strip is placed perpendicularly across the ankle, and the second is placed perpendicularly to the first strip, parallel to the skin incision and to the right of the leg. Press the 2 strips down lightly on the block of blu-tack to secure the leg, but taking care not to press down too tightly to obstruct blood flow ().
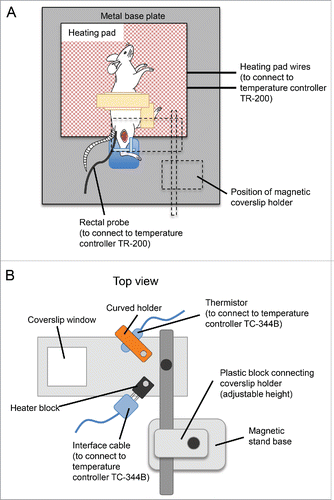
Figure 3. Schematic diagram of the setup for the tibialis anterior muscle intravital multiphoton imaging. (A) Schematic diagram of the imaging stage, showing the position of the mouse on the heating pad, with the right tibia in the blu-tack mold. Position of the coverslip holder is outlined in dotted lines. (B) Top view of the coverslip holder. Positions of the thermistor and interface cable, to be connected to the curved holder and heater block, respectively, are shown.
Finally, we tape the free left leg down on the heating pad to secure it (). The breathing movements of the mouse may cause the imaging area to be unstable. To minimize vibrations, we cut a 12 cm × 2.5 cm strip of c-fold paper towel, fold it in half and place it over the mouse like a blanket, adhering it to the heating pad with masking tape ().
To image the outer connective tissue, a ring of vacuum grease can be applied around the window in the skin () and filled with a drop of PBS, and a coverslip (attached on the coverslip support holder) is placed over the well lightly but firmly (). Ensure that the well of vacuum grease is sealed on all sides to prevent PBS from leaking out during the imaging session and causing the tissue to dry up. Pressing the coverslip too hard will stop the blood flow and affect the imaging results. The tissue is now ready to be imaged.
To image the skeletal muscle and capillaries, the outer connective tissue should be removed. We place the mouse with the stage under a dissecting microscope. A major connective tissue runs as a vertical white line down the muscle tissue (arrow in ), and there is a lower density of blood vessels on the right side of this connective tissue. To minimize bleeding during the preparation, we remove the outer connective tissue layer on the right side of this line. Starting from the top of the incision, use fine forceps to lift up the thin layer of outer connective tissue gently, and make a small cut with the dissecting spring scissors. Continue to cut a small window in the connective tissue to expose an area of the muscle fibers large enough for imaging, lifting the connective tissue gently as you proceed. Take extra care not to pull hard on the tissue before cutting with the scissors, as this will injure the muscle. The outer connective tissue is a reflective surface, and a clean removal of this outer layer will reveal a shiny surface of the muscle tissue with the associated striations visible (). We can then proceed to apply the ring of vacuum grease, PBS and coverslip as previously described ().
Note: Application of reagents directly on muscle tissue
Reagents such as inflammatory agents or chemoattractants can be directly applied on the skeletal muscle. A ring of grease should be applied to the muscle first, to create a reservoir for the reagent. The well should hold about 100 μl of reagent.
Move the imaging stage to the microscope chamber. For the coverslip support holder, insert the thermistor for temperature monitor in the curved holder, and connect the interface cable for temperature control to the resistive heater block (). Insert the rectal probe into the mouse rectum gently (). Connect the heating pad and the rectal probe to the temperature controller, and hence the heating pad will be regulated by the feedback rectal probe (). We recommend maintaining the temperature of the heating pad and coverslip holder at 36°C and 32°C, respectively, to prevent the overheating of the mouse and muscle tissue.
We also typically inject in Evans Blue intravenously into the mouse by retroorbital injection to allow visualization of the vasculature. Other fluorescent dyes or other reagents can also be injected as desired, via retroorbital or tail vein injection. We usually image 15 minutes at baseline to ensure that there is no preparation-related injury. If Evans Blue dye had been injected to label blood vessels, any damage to blood vessels during preparation of mouse for imaging will show up as patches of blue corresponding to fluid leakage.
Lastly, add a drop of water with a transfer pipette on the coverslip, and gently lower the water-dipping objective lens on the water. Focus the specimen using the eyepiece and choose an area of interest as desired. Commence two-photon intravital imaging according to the manufacturer's instructions.
Demonstration of tibialis anterior muscle architecture and components
We utilize the LysM-GFP (Lyz2GFP/+) Citation15 and the ROSA-mT/mG (Gt(ROSA26)ACTB-tdTomato-EGFP) Citation16 mouse to demonstrate the different layers and components of the skeletal muscle. The connective tissue produces second harmonic generation (SHG) signals that can be easily captured at half the excitation wavelength. In the LysM-GFP mouse, the neutrophils are GFP-high, while the monocytes and macrophages are GFP-intermediate. For our imaging, we use an excitation wavelength of 950 nm. The microscope filters used are 495 long-pass (LP; Chroma, cat. no. T495LPXR) and 560 LP (Chroma, cat. no. T560LPXR), and the mirror sets used are 475/42 band-pass (BP; Semrock, cat. no. FF01-475/42-25) and 510/20 BP (Semrock, cat. no. FF03-510/20-25).
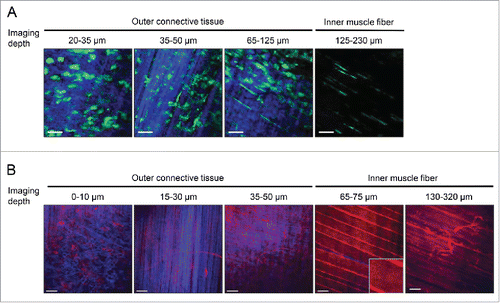
We typically observe 3 layers of outer connective tissue if this outer layer is not surgically removed during the preparation (). The top-most layer consists of a mesh-like network of connective tissue (panel 1), the second layer shows well-defined vertical striations (panel 2), while the third layer shows horizontal striations that run perpendicular to the second layer (panel 3). Muscle tissue macrophages reside in these layers of outer connective tissue. The inner muscle fiber deeper down has no SHG, and has no muscle tissue macrophages present. Instead, we observe the perivascular macrophages that line the capillaries within the muscle fiber layer (panel 4). The outer connective tissue layer is typically about 100 μm, hence, to capture better signal from the inner muscle fiber and image the capillaries, we surgically remove the outer connective tissue.
Figure 4. Visualization of outer connective tissue layers and inner muscle fiber/capillaries. (A) Imaging of the outer connective tissue layers and the inner muscle fiber layer in a LysM-GFP mouse. Muscle macrophages (green) are present in the outer connective tissue layers. SHG shows striations of the connective tissue. Three layers of connective tissues (blue) can be typically captured: Top-most layer has a mesh network, second layer has distinct vertical striations, and third layer has horizontal striations perpendicular to the second layer. Inner muscle fiber has minimal SHG and perivascular macrophages (green) can be seen lining the blood vessels. (B) Imaging of the outer connective tissue layers and the inner muscle fiber layer in a ROSA-mT/mG mouse. Three layers of connective tissues (blue) similar to (A) are shown: Top-most layer has a mesh network, second layer has distinct vertical striations, and third layer has horizontal striations perpendicular to the second layer. Muscle macrophages and blood vessels can be seen (red) in the outer connective tissue layers. Parallel muscle fibers are seen in the inner muscle fiber layer. Striations of the muscle can be seen (zoomed in; inset). Muscle capillaries are present deeper in the muscle fiber layer. Excitation: 950 nm using a Ti:Sapphire laser. Scale bar: 50 μm. Step size: 2–4 μm.
Using the LysM-GFP mouse, if there was mechanical damage during the muscle preparation, neutrophils will accumulate at the injury site quickly, and clusters of bright green cells will be observed in the inner muscle fiber layer where the capillaries are situated. This does not represent the baseline physiology of a non-inflamed muscle tissue, hence the muscle preparation should be repeated with another mouse.
Next, using the ROSA-mT/mG mouse, the ubiquitous ROSA promoter will drive membrane expression of tdTomato fluorescent protein in all cells. For our imaging, we use an excitation wavelength of 950 nm. The microscope filters used are 560 LP (Chroma, cat. no. T560LPXR) and 640 LP (Chroma, cat. no. 640 DCLP), and the mirror sets used are 475/42 BP (Semrock, cat. no. FF01-475/42-25) and 579/34 BP (Semrock, cat. no. FF01-579/34-25). In the ROSA-mT/mG mouse, we demonstrate the 3 layers of connective tissue as well (). Muscle macrophages and blood vessels show expression of the tdTomato fluorescent reporter. Deeper in the muscle tissue, we observe parallel muscle fibers along the long axis of the tibialis muscle (). The internal ultrastructure of repeating sarcomeres is also visible along skeletal muscle fibers (). Finally, deeper in the inner muscle fiber, we can observe the capillaries and post-capillary venules using the ROSA-mT/mG mouse ().
Demonstration of intravenous delivery of agents via jugular catheter
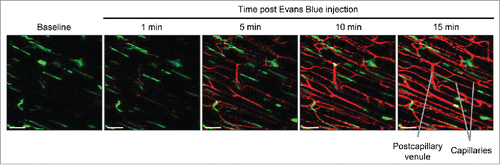
We perform the jugular vein administration of Evans Blue dye intravenously in a LysM-GFP mouse, to demonstrate the rapid labeling of the vasculature. The left jugular vein of the mouse is cannulated and 50 μg Evans Blue dye (Sigma-Aldrich) dissolved in 50 μl saline is introduced very slowly using the syringe attached to the catheter after 10 min of baseline recording. The first labeling of the vasculature is visible within a minute post-introduction (, supplementary video 1). For our imaging, we use an excitation wavelength of 950 nm. The microscope filters used are 495 LP (Chroma, cat. no. T495LPXR) and 560 LP (Chroma, cat. no. T560LPXR), and the mirror sets used are 475/42 BP (Semrock, cat. no. FF01-475/42-25), 510/20 BP (Semrock, cat. no. FF03-510/20-25) and 665/40 BP (Chroma, cat. no. ET665/40M-2P). Images were captured at 1 frame per min, and videos were recorded at 6 frames per second.
Figure 5. Evans Blue intravenous delivery using jugular vein catheter. Snapshot images of the inner muscle fiber layer from in vivo imaging during a slow intravenous delivery of Evans Blue via the jugular vein catheter into a LysM-GFP mouse. Evans Blue is slowly delivered to the mouse at time t=10 min. Snapshot images are shown at 1 min, 5 min, 10 min and 15 min after the start of Evans Blue introduction. Perivascular macrophages (green) line the blood vessels (red). Excitation: 950 nm using a Ti:Sapphire laser. Scale bar: 50 μm.
Using the jugular vein administration, the muscle can be imaged simultaneously while injecting agents into the mouse to observe the downstream effects. For labeling of the vasculature, we recommend using Evans Blue, compared to other reagents such as fluorescent dextran or lectin, as Evans Blue has a sharp emission peak at 680 nm. This will result in minimal spectral overlap with other commonly used fluorophores such as GFP and RFP to allow for co-visualization.
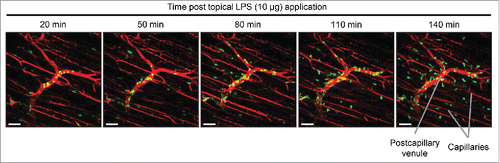
Next, we demonstrate the tibialis anterior skeletal muscle imaging procedure by utilizing the LysM-GFP mouse again, to visualize the circulating immune cells in the muscle capillaries (labeled with Evans Blue) and their response to an inflammatory agent LPS topically applied to the muscle tissue. We applied 10 μg of LPS directly on the exposed muscle tissue of a LysM-GFP mouse, set up the imaging immediately, and imaged the muscle after LPS application. A typical time duration from reagent application to the start of an imaging session is about 10–20 min. In , we show the first image taken 20 min after LPS application. The muscle is imaged for 2 hours post-treatment, and we observe the extravasation of neutrophils (GFP-high) from the post-capillary venules (, supplementary video 2), at an average speed of about 9.8 μm/min (N = 35). Images were captured at 1 frame per min, and videos were recorded at 6 frames per second.
Figure 6. In vivo imaging of inflammatory agent on muscle tissue. Snapshot images of the inner muscle fiber layer from in vivo imaging after topical application of LPS (10 μg) on the tibialis anterior skeletal muscle of a LysM-GFP mouse. GFP-high myeloid cells (mostly neutrophils) extravasate from the blood vessels (red; labeled via i.v. injection of Evans Blue). Excitation: 950 nm using a Ti:Sapphire laser. Scale bar: 50 μm.
Discussion
We provide a detailed protocol of the preparation of the mouse for intravital imaging of the tibialis anterior skeletal muscle, as well as the steps for jugular vein catheterization to allow for infusion of compounds during imaging. We demonstrate the use of tibialis anterior muscle imaging to visualize in vivo transmigration of immune cells in muscle capillaries, in response to LPS stimulation (, Supplementary video 2). We also demonstrated the use of jugular vein catheterization by injection of Evans Blue dye (, Supplementary video 1).
Selection of imaging areas in the tibialis skeletal muscle
The skeletal muscle is dense with capillaries and is hence an apt organ for visualizing immune-vasculature-endothelial interactions due to the vast and dense capillary bed. We typically image an area that includes capillaries and a post-capillary venule to enable us to visualize leukocyte rolling and extravasation (Supplementary Fig. 1). The muscle tissue is striated (), and the capillaries form a high-density meshwork between and parallel to the muscle fibers. Arterioles and venules are larger/broader than capillaries, which are one cell thick. Blood flow is faster in the arterioles than the postcapillary venules. Using a fluorescent reporter mouse of any leukocyte population, one will be able to visualize leukocytes zoom by fast and thus appear as “streaks,” while in venules, one will observe leukocyte rolling. Post-capillary venules are typically Y-shaped blood vessels, from which capillaries (thinner vessels) converge upon. Blood flow is also seen directed from the capillaries into the venules.
Anesthesia monitoring
Preparation of the animal for jugular catheterization and setup of the muscle imaging, together with the time-lapse imaging experiment can last over 1–2 hours, and may extend longer. We always visually inspect the anesthesia condition of the mouse during the whole process and re-anesthetize the mouse as necessary. Our first dose of 150 mg/kg ketamine and 10 mg/kg xylazine via intraperitoneal injection typically lasts one hour, and we re-anesthetize a quarter dose subsequently for every half hour via subcutaneous injection to keep the mouse under deep anesthesia. For better monitoring of the physiological state of the mouse and to minimize adverse effects of overdosing on anesthetics, electrocardiograms, pulse oximeters or microcapnographs can be used. Additionally, for longer imaging sessions, investigators may want to consider other anesthetics such as inhalation of isoflurane or injection of avertin. Citation17
Continuous imaging
We have typically imaged the muscle for about 2–3 hours. However, since the tibialis skeletal muscle preparation is minimally invasive, the imaging session can last longer if required. The investigator will need to carefully monitor the mouse condition, ensure that the fluid in the grease well is not dried up, and constantly replenish the immersion water droplet for the objective lens.
Application of the tibialis anterior muscle imaging model
Skeletal muscle imaging can be further applied to visualize the processes of muscle contraction. Since muscle contraction requires high amount of energy consumption, the muscle tissue is rich in mitochondria and this imaging setup can be used to study mitochondria biology. If the outer connective tissue is not removed during preparation, this setup can be used to image the outer layer of connective tissue.
Vaccines are often administered intramuscularly. Hence, preclinical trials of vaccines and other drug compounds can utilize this imaging setup to examine the biological effect and biodistribution of a new vaccine through direct visualization of the muscle tissue.
Comparison with other muscle tissue
The other muscle tissue commonly used for intravital imaging is the cremaster muscle that covers the testicles. Preparation of the tibialis anterior muscle for intravital imaging is simple compared to the cremaster muscle. The skeletal muscle is exposed through a simple incision in the skin. Mounting the tissue straightaway will allow the imaging of the connective tissue, and careful removal of the outer connective tissue with surgical scissors will expose the muscle tissue, allowing imaging of muscle fibers and blood capillaries. In contrast, imaging of the cremaster muscle involves more surgical manipulations to the muscle itself. The muscle is exposed, detached from the testicles, and spread radially as a flat sheet. This procedure of spreading the cremaster muscle tissue may induce significant inflammation. outlines the comparison of the cremaster muscle and tibialis muscle.
Table 1. Comparison of cremaster muscle and tibialis muscle for intravital imaging.
Materials
Reagents required:
Experimental mice
PBS
Saline
Ketamine
Xylazine
Evans Blue (or other vessel-labeling agent)
Chemoattractant (if required)
Water
Equipment required:
Insulin syringe with needle (BD, cat. no. 328838)
Transfer pipettes (Biologix Research Company, cat. no. 30–0135)
3M paper masking tape (3M, cat. no. 2214)
Deckglaser microscope cover glass (22 mm × 32 mm; Superior Marienfeld Laboratory Glassware, Paul Marienfeld, cat. no. 0101112)
Medical cotton-tipped applicators (Puritan Medical Products Company, cat. no. 806-WC)
Surgical microscope (Leica, model M651 MSD)
Custom-built muscle-imaging stage platform (see EQUIPMENT SETUP
for details)
McPherson-Vannas Micro Dissecting Spring Scissors (Roboz, cat. no. RS-5600)
Dumont Tweezers (Roboz, cat. no. RS-5047)
Extra Fine Micro Dissecting Scissors (Roboz, cat. no. RS-5880)
Curved splinter forceps (Aesculap, B. Braun Melsungen, cat. no. BD314R)
Braun cruZer6 precision trimmer (Braun)
1 mL Luer Slip Tuberculin Syringe (BD, cat. no. 302100)
Re-usable adhesive (Blu-tack)
Polyethylene tubing for catheter (Microtube Extrusions, cat. no. PE6128)
BD PrecisionGlide 30G × 1/2 in. needle (BD, cat. no. 305106)
Black braided silk surgical suture (Roboz, cat. no. SUT-15–1)
C-fold towels (Kimberly-Clark, cat. no. 20311)
Dow Corning ® high vacuum grease
18.5 × 20.5 cm plastic board
Equipment setup:
Magnets (4.0 × 2.0 × 1.5 cm)
Metal base plate (19.5 × 19.5 cm)
Styrofoam sponge (12.5 × 10.0 cm or cut to the size of available heating pad)
Heating pad (Fine Science Tools, cat. no. 21060–00)
Curved holder, for positioning the control thermistor
Interface cable CC-28 with DIN connector and thermistors, one for the (temperature control and the other for the temperature monitor ((Warner Instruments (Harvard Apparatus), cat. no. 640106)
Resistive heater block RH-2 (Warner Instruments (Harvard Apparatus), (cat. no. 640274) (
Temperature controller TC-344B for the ear platform (Warner Instruments ((Harvard Apparatus), cat. no. 640101)
Temperature controller TR-200 for mouse heating pad (Fine Science Tools, (cat. no. 21052–00) (
Power supply for TR-200 (Fine Science Tools, cat. no. 21051–00)
Animal rectal probe (Fine Science Tools, cat. no. 21060–01)
C-fold towels (Kimberly-Clark, cat. no. 20311)
Coverslip support holder: Plastic base (5.0 × 3.5 × 0.5 cm) with magnet embedded. Base is the support for a plastic tube that is 8.5 cm in length, 1.0 cm in circumference. Attach the resistive heater block and the curved holder for the control thermistor on the coverslip holder, using mounting screws.
Intravital imaging microscope
Mice were imaged using the TriM Scope II single-beam 2-photon upright microscope (LaVision BioTec). The microscope is equipped with a tunable (680–1080 nm) Coherent Chameleon Ultra II One Box Ti:Sapphire laser (3.3 W at 800 nm; pulse length of 140 fs, 80 MHz repetition rate; Coherent) and the following photomultiplier modules: blue (quantum efficiency (QE) = 17% at 400 nm; Hamamatsu, cat. no. H 6780–01), green/red (QE = 15% at 630 nm; Hamamatsu, cat no. H6780–20), high sensitivity module (QE = 40% at 550 nm; Hamamatsu, cat. no. H7422–40). Imaging was performed using a 20x water-dipping objective (NA = 1.0; XLUMPLFLN20xW, Olympus), and AHS LAMP 12 V/100 W halogen lamp was used for wide-field epifluorescence.
Mice
Mice were bred and maintained in the A*STAR Biological Resource Center (BRC) mouse facility. All experiments were approved by and performed according to the BRC Institutional Animal Care and Use Committee (IACUC), in compliance with the guidelines of the Agri-Food and Veterinary Authority (AVA) and the National Advisory Committee for Laboratory Animal Research (NACLAR) of Singapore.
Image analysis
Images were processed into time series movies and analyzed using IMARIS image analysis software (Bitplane). The plugin “Correct 3D Drift” in Fiji program was used for drift correction of images acquired by time-lapse imaging. Citation18
Abbreviations
| BP | = | band-pass |
| LP | = | long-pass |
| LPS | = | lipopolysaccharide |
| SHG | = | second harmonic generation |
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Supplemental_Video_2.mov
Download QuickTime Video (19.9 MB)Supplemental_Video_1.mov
Download QuickTime Video (2.8 MB)1156272_Supplemental_Material.zip
Download Zip (2 MB)Acknowledgments
We would like to thank Prof. Thomas Graf for kindly providing LysM-GFP mice, and Maximilien Evrard and Jackson L.Y. Li for technical help and comments on the manuscript.
Funding
This work was supported by the Singapore Immunology Network (SIgN), Agency for Science, Technology and Research of Singapore (A*STAR).
References
- Hoppeler H, Mathieu O, Weibel ER, Krauer R, Lindstedt SL, Taylor CR. Design of the mammalian respiratory system. VIII. Capillaries in skeletal muscles. Respir Physiol 1981; 44:129-50; PMID:7232883; http://dx.doi.org/10.1016/0034-5687(81)90080-3
- Welling TH, Davidson BL, Zelenock JA, Stanley JC, Gordon D, Roessler BJ, Messina LM. Systemic delivery of the interleukin-1 receptor antagonist protein using a new strategy of direct adenoviral-mediated gene transfer to skeletal muscle capillary endothelium in the isolated rat hindlimb. Hum Gene Ther 1996; 7:1795-802; PMID:8894671; http://dx.doi.org/10.1089/hum.1996.7.15-1795
- Bagher P, Segal SS. The mouse cremaster muscle preparation for intravital imaging of the microcirculation. J Vis Exp 2011; 52:2874; PMID:21694690
- Akimoto T, Sorg BS, Yan Z. Real-time imaging of peroxisome proliferator-activated receptor-gamma coactivator-1alpha promoter activity in skeletal muscles of living mice. Am J Physiol Cell Physiol 2004; 287:C790-6; PMID:15151904; http://dx.doi.org/10.1152/ajpcell.00425.2003
- Rudolf R, Mongillo M, Magalhães PJ, Pozzan T. In vivo monitoring of Ca(2+) uptake into mitochondria of mouse skeletal muscle during contraction. J Cell Biol 2004; 166:527-36; PMID:15314066; http://dx.doi.org/10.1083/jcb.200403102
- Devi S, Wang Y, Chew WK, Lima R, A-González N, Mattar CNZ, Chong SZ, Schlitzer A, Bakocevic N, Chew S, et al. Neutrophil mobilization via plerixafor-mediated CXCR4 inhibition arises from lung demargination and blockade of neutrophil homing to the bone marrow. J Exp Med 2013; 210:2321-36; PMID:24081949; http://dx.doi.org/10.1084/jem.20130056
- Hoppeler H, Fluck M. Plasticity of skeletal muscle mitochondria: structure and function. Med Sci Sports Exerc 2003; 35:95-104; PMID:12544642; http://dx.doi.org/10.1097/00005768-200301000-00016
- Rothstein EC, Carroll S, Combs CA, Jobsis PD, Balaban RS. Skeletal muscle NAD(P)H two-photon fluorescence microscopy in vivo: topology and optical inner filters. Biophys J 2005; 88:2165-76; PMID:15596503; http://dx.doi.org/10.1529/biophysj.104.053165
- Glancy B, Hsu L-Y, Dao L, Bakalar M, French S, Chess DJ, Taylor JL, Picard M, Aponte A, Daniels MP, et al. In vivo microscopy reveals extensive embedding of capillaries within the sarcolemma of skeletal muscle fibers. Microcirculation 2014; 21:131-47; PMID:25279425; http://dx.doi.org/10.1111/micc.12098
- Prakash MD, Munoz MA, Jain R, Tong PL, Koskinen A, Regner M, Kleifeld O, Ho B, Olson M, Turner SJ, et al. Granzyme B Promotes Cytotoxic Lymphocyte Transmigration via Basement Membrane Remodeling. Immunity 2014; 41:960-72; PMID:25526309; http://dx.doi.org/10.1016/j.immuni.2014.11.012
- Azcutia V, Routledge M, Williams MR, Newton G, Frazier WA, Manica A, Croce KJ, Parkos CA, Schmider AB, Turman MV, et al. CD47 plays a critical role in T-cell recruitment by regulation of LFA-1 and VLA-4 integrin adhesive functions. Mol Biol Cell 2013; 24:3358-68; PMID:24006483; http://dx.doi.org/10.1091/mbc.E13-01-0063
- Schymeinsky J, Gerstl R, Mannigel I, Niedung K, Frommhold D, Panthel K, Heesemann J, Sixt M, Quast T, Kolanus W, et al. A fundamental role of mAbp1 in neutrophils: impact on β2 integrin–mediated phagocytosis and adhesion in vivo. Blood 2009; 114:4209-20; PMID:19638624; http://dx.doi.org/10.1182/blood-2009-02-206169
- Stokol T, O'Donnell P, Xiao L, Knight S, Stavrakis G, Botto M, Andrian von UH, Mayadas TN. C1q governs deposition of circulating immune complexes and leukocyte Fcgamma receptors mediate subsequent neutrophil recruitment. J Exp Med 2004; 200:835-46; PMID:15466618; http://dx.doi.org/10.1084/jem.20040501
- Sperandio M, Thatte A, Foy D, Ellies LG, Marth JD, Ley K. Severe impairment of leukocyte rolling in venules of core 2 glucosaminyltransferase-deficient mice. Blood 2001; 97:3812-9; PMID:11389021; http://dx.doi.org/10.1182/blood.V97.12.3812
- Faust N, Varas F, Kelly LM, Heck S, Graf T. Insertion of enhanced green fluorescent protein into the lysozyme gene creates mice with green fluorescent granulocytes and macrophages. Blood 2000; 96:719-26; PMID:10887140
- Muzumdar MD, Tasic B, Miyamichi K, Li L, Luo L. A global double‐fluorescent Cre reporter mouse. Genesis 2007; 45:593-605; PMID:17868096; http://dx.doi.org/10.1002/dvg.20335
- Ewald AJ, Werb Z, Egeblad M. Monitoring of vital signs for long-term survival of mice under anesthesia. Cold Spring Harb Protoc 2011; 2011:pdb.prot5563
- Parslow A, Cardona A, Bryson-Richardson RJ. Sample Drift Correction Following 4D Confocal Time-lapse Imaging. J Vis Exp 2014; 86:e51086-6