
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Late leaf spot and groundnut rosette are major constraints to groundnut cultivation, especially in the second crop cycle in central Côte d’Ivoire. This study was initiated to improve peanut production. It was conducted on 22 peanut accessions using a randomized complete block design with three (03) replications. In each elementary plot, agromorphological, and health parameters were assessed on ten plants per accession. A symptom rating scale was used to assess disease severity. The results showed that the agromorphological parameters were higher in the first cycle than in the second. Accessions ARA42, ARA60, ARA56, and ARA51, moderately resistant or tolerant to groundnut rosette and susceptible to late leaf spot, gave the best seed yields in the second cycle, which varied between 1.42 and 1.59 tons per hectare (t/ha). Those highly susceptible to these parasitic diseases (ARA55, ARA40, ARA39, ARA54, ARA57, and ARA53) gave the lowest yields ranging from 0.131 to 0.996 t/ha over the same period. Groundnut rosette and late leaf spot attacks were less severe in the first crop cycle than in the second, with mean severity ranging from 36.5 to 51.5% and 24.9 to 55.5%, respectively. Accessions ARA42, ARA60, ARA56, and ARA51 should be the subject of a rigorous selection program and made available to growers. As a result, yield losses during the second crop cycle, from July to October, will be reduced, and the use of synthetic chemical pesticides in peanut cultivation will be limited.
Public Interest Statement
The manuscript reveals the biotic constraints linked to groundnut production in the second crop cycle between July and October. This document is important for public authorities, development and groundnut sector stakeholders and small-scale producers who make a living from this activity. The article shows that production losses in the second crop cycle are more than 60%. But these losses can be reduced or mitigated by using resistant or tolerant genotypes. To this end, this paper presents resistant or tolerant groundnut varieties that can be used in the second crop cycle to limit production losses. Given the importance of groundnuts in cropping systems, in the agri-food industry, for human and animal consumption, for medicinal purposes and as a source of income for small-scale producers, this article provides an opportunity to produce groundnuts over both cropping cycles (March to June and July to October).
Reviewing Editor:
Introduction
Peanut (Arachis hypogaea L.) is an oilseed crop grown in more than 100 countries in tropical, subtropical, and warm temperate regions (Guchi et al., Citation2014). It is the 13th most important food crop and the 4th most important oilseed crop worldwide (Reddy et al., Citation2011). It is produced in much of sub-Saharan Africa for commercial and domestic consumption (Christie et al., 2015). It is consumed as seeds after the pods have been hulled, after the seeds have been crushed, or in more or less elaborate forms (butter, pasta, flour, various confectionery products). The seeds are rich in protein (25%) and edible oil (50%), making them a highly prized foodstuff. In addition, peanut by-products, notably oilcakes, and tops, are used in animal feed (Marloire, Citation2019). It is also present in cropping systems thanks to its nodules, which are capable of fixing atmospheric nitrogen, which is beneficial for soil fertilization (Coulibaly et al., Citation2021).
In Côte d’Ivoire, peanuts are produced mainly in the northern and central regions. Annual national production is estimated at 112,698 tons in 2019 (FAOSTAT, Citation2020). Current production is insufficient to cover the population’s annual needs, estimated at 150,000 tons. To meet these needs, national production must be improved, particularly by carrying out two crop cycles (Ngoyi et al., Citation2015). However, in second-cycle conditions, peanut cultivation is more prone to diseases and pests (Gadji et al., Citation2021). Indeed, in second-cycle conditions, peanuts, once emerged, are subject to severe attacks during vegetation by fungal and viral diseases whose damage causes a drop in yield (Blizoua Bi & Djan, Citation1995; Gadji et al., Citation2021; N’Gbesso et al., Citation2020) and a deterioration in the biological, technological and nutritional quality of the products (Manizan et al., Citation2018).
Among the diseases that reduce producers’ efforts, two are more harmful to peanuts and are capable of causing significant production losses in the West African zone. These are late leaf spot and groundnut rosette (Okello et al., Citation2010). The main fungal leaf disease of cropped peanut, late leaf spot, causes yield losses of up to 40% during prolonged periods of high relative humidity and at daily temperatures of 16 °C to 25 °C (Zinsou et al., Citation2019). There is also groundnut rosette disease (virosis), which, in the event of an epidemic, leads to a total loss of production if the year’s climatic conditions are conducive to early infestation (Schilling et al., Citation1996).
Several control methods, including the use of preventive cultural techniques and the application of plant protection products, are used to reduce the incidence of these diseases. However, these measures are only used to a limited extent due to the difficulties encountered when implementing them and the danger to the environment and consumer health posed by the misuse of chemical pesticides (Van Lenteren et al., Citation2018). As a result, biological methods have been developed to combat these diseases. Such methods include varietal control using resistant varieties to prevent the disease in all circumstances. Schilling et al. (Citation1996) have shown satisfactory results using resistant varieties in areas where peanut rosette and leaf spot are endemic. Based on this finding, selecting leaf spot and groundnut rosette-tolerant peanuts is essential, as it appears to be an effective solution for improving yields in second-cycle conditions. This study aims to select high-yielding peanut accessions tolerant or resistant to late leaf spot and groundnut rosette diseases to limit production losses in the second crop cycle.
Materials and methods
Study area
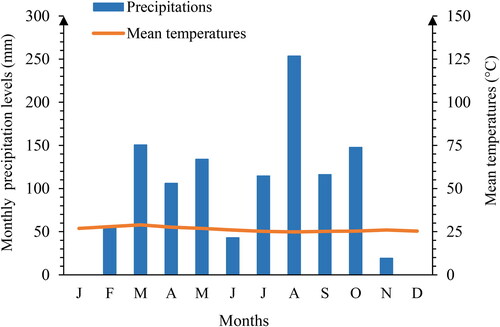
The trial was conducted at the food crop research station of the Centre National de Recherche Agronomique (SRCV/CNRA) located at Bouaké in central Côte d’Ivoire at latitude 7°40’42.94” N, longitude 5°5’59.37” W and altitude 401 m. Precipitation recorded during the study period, from March to June 2019, ranged from 150.5 to 42.9 mm and between July and October from 114.6 to 147.7 mm, with temperatures varying between 25.2 and 28.96 °C () and relative humidity between 62 and 85%.
Figure 1. Umbrothermal curve (data source: CNRA, 2019).
Plant material
Twenty-two (22) peanut accessions were tested. These accessions were coded as follows: ARA for groundnut, the following digits represent the registration number in the CNRA files, the places of provenance and the home country ().
Table 1. Peanut accessions used as seed and their origin of acquisition.
Experimental design
The trial was conducted using a randomized complete block design with three replications. The plot was 34 m long and 20 m wide with a 1 m border on each side. The total surface area of the trial was 792 m2. Each block (replication) consisted of 8 elementary plots. The individual plots and blocks were 2 m apart. The dimensions of each elementary plot were 4 m in length and 2 m in width, giving a surface area of 8 m2. Each elementary plot consisted of three (3) rows of seedlings corresponding to three (3) distinct accessions. Eleven (11) seedlings were grown in each row, giving 33 plants per elementary plot. These plants were spaced 0.4 m apart along the row and 1 m between rows over the two growing cycles.
Agronomic parameters observed
Observations were made on ten plants taken randomly from each line, from 3rd to 12th plants, per accession on each elementary plot per the method described by Gadji et al. (Citation2021). These observations covered growth parameters, phenological stages, and yield components. Growth parameters included plant height (HP) and crown diameter (DC). HP and DC were measured per accession on each elementary plot using a graduated ruler and a caliper one day before harvest. As for the various phenological stages, emergence, flowering, and ripening times were recorded when 50% of the plants had reached the stage indicated. The yield components determined were the percentage of pods with one seed, two seeds, three seeds, and the weight of 100 dry seeds. The net yield was determined using the EquationEquation (1)(1)
(1) below:
(1)
(1)
The percentage of 1-seed, 2-seed, and 3-seed pods was determined for each accession from the pods of 5 randomly selected plants;
One hundred (100) dried groundnut seeds were weighed per accession for each elementary plot to determine their weight.
Disease’s assessment
The late leaf spot severity index was assessed on ten plants per accession on each elementary plot, 75 days after sowing (JAS), using the rating scale described by Subrahmanyam et al. This scale evaluates the percentage of leaf area infected by the disease and varies from 1 to 9 where 1 = 0%, 2 = 1%–5%, 3 = 6%–10%, 4 = 11%–20%, 5 = 21%–30%, 6 = 31%–40%, 7 = 41%–60%, 8 = 61%–80%, 9 = 81%–100%. The ten plants were randomly selected in a row, as described above.
In the case of groundnut rosette, the severity assessment on the plants was based on the symptom intensity scale described by James (Citation1974), to which scores were assigned using the method of Olorunju (Citation1991). Thus, the severity index rating scale was defined as follows:
0: healthy plant/no visible symptoms (0%)
1: Ring spots/presence of symptoms but no stunting (1% to 20%)
2: Dense grouping of stunted leaves at the terminal (21% to 40%)
3: Chlorosis/mosaic with 30% of plant volume stunted (41% to 60%)
4: Extreme stunting with 30 to 70% of plant volume stunted (61% to 80%)
5: Death of terminal leaves with more than 70% of plant volume stunted (81% to 100%).
The mean severity index was calculated according to the following EquationEquation (2)(2)
(2) :
(2)
(2)
Where ni = sum of individual indices as a percentage 75 days after sowing, N = total number of plants observed per accession and elementary plot.
Statistical analysis of data
The data collected were analyzed using Statistica 7.1 for analysis of variance and XLSTAT 2018 version Pro 7.5 for principal component analysis (PCA) and hierarchical ascending classification (HAC). One-way analyses of variance (ANOVA 1) were performed to obtain the means of the various agromorphological and health parameters for each accession. Where a significant difference was found, the LSD Fisher test at the 5% threshold was used to classify the averages into homogeneous groups. Principal component analysis (PCA) was used to characterize accessions and study correlations between disease severity and agromorphological parameters of groundnut accessions in the first and second cropping cycles. Hierarchical ascending classification (HAC) highlighted the various homogeneous groups within the peanut collection. These methods were used to elucidate the nature and structure of the agromorphological variability of the accessions studied.
Results
Phenological stages
The results for the different phenological stages are shown in . Analysis of variance revealed significant differences (p < 0.05) between peanut accessions concerning the parameters studied. In general, the accessions recorded, on average, earlier emergence (6 JAS) and maturation (107 JAS) dates in the second crop cycle than in the first, except for flowering time. However, it should be noted that the earliest accessions for all crop cycles were ARA39, ARA40, and ARA54, and the latest were ARA42, ARA44, ARA45, ARA59, and ARA60 for flowering and ripening times. The dispersion of the values measured around the mean ranged from 21.42 to 1.58 and decreased from emergence to ripening time.
Table 2. Phenological times (emergence, flowering, and ripening) for peanut accessions.
Mean severity of groundnut rosette and late leaf spot
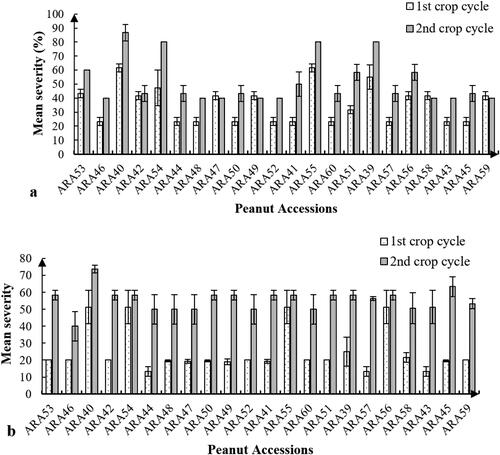
The evolution of the severity of rosette and late leaf spot symptoms over the two crop cycles is illustrated in . Disease severity varied according to the accessions, with higher severity in the second crop cycle than in the first. Statistical analyses revealed significant differences (F(21, 44) ≥ 28.650 with P = 0.001) between the accessions studied regarding groundnut rosette severity and highly significant differences (F (21, 44) ≥ 3.744 with P = 0.0001) concerning the severity of late leaf spot, whatever the crop cycle. Statistical data showed that the first crop cycle was the least attacked, with an average severity between 24.9 and 35.6%, compared to the second crop cycle, with an average severity between 51.5 and 55.5% for the peanut rosette () and late leaf spot (), respectively. However, most of the peanut accessions evaluated showed tolerance to groundnut rosette, except for six accessions, ARA39, ARA40, ARA51, ARA54, ARA55, and ARA56. These same accessions were also susceptible to late leaf spots in the first and second crop cycles.
Figure 2. Mean severity of groundnut rosette (a) and late leaf spot; (b) over the two crop cycles.
Yield and its components
presents the results relating to the number of pods with one (01), two (02), and three (03) seeds, 100-seed weight, and seed yield obtained during the two cropping cycles. The results show that the number of one-seeded and two-seeded pods was lower in the second crop cycle than in the first, while it was almost zero for three-seeded pods. Statistical analyses of variance revealed significant differences (F(21, 44) ≥ 2.519 avec P ≤ 0.0048) between peanut accessions for these parameters, except for the first cycle for yield and both cycles for 100-seed weight. The best seed yields were obtained with accessions ARA42 (2.72 and 1.594 t/ha), ARA44 (3.04 and 1.22 t/ha), ARA45 (4.8 and 1.21 t/ha), ARA50 (2.9 and 1.23 t/ha), ARA58 (2.6 and 1.4 t/ha), ARA59 (5.05 and 1.3 t/ha), and ARA60 (2.97 and 1.5 t/ha) over the two crop cycles. Statistical analysis showed no significant difference (p > 0.05) between these accessions regarding seed yield. On the other hand, the lowest seed yields were recorded with accessions ARA39 (1.34 and 0.23 t/ha) and ARA57 (1.33 and 0.71 t/ha). Yield losses in the second cycle due to diseases were 62.1%.
Table 3. Yield and its components over the two crop cycles.
Agromorphological diversity of the studied accessions
Structuring and characterization of accessions studied in the first growing cycle
PCA was used to group the most discriminating descriptors according to their correlation. The first two axes (F1 and F2) represent 49.88% of the total variability and are characterized by production, health status, and ripening time. Axis 1 (F1) represents 29.51% of the total variability and is characterized by the production and health status of the accessions. The eigenvalues obtained for the production variables show that flowering time (FD), number of pods (NG), number of seeds (Ngr), and seed yield (Rgr) are positively correlated (). This correlation shows seed yield is related to the number of pods and seeds per plant.
Figure 3. Correlation circle (a) and dispersion of accessions; (b) on the F1 and F2 axes in the first crop cycle.
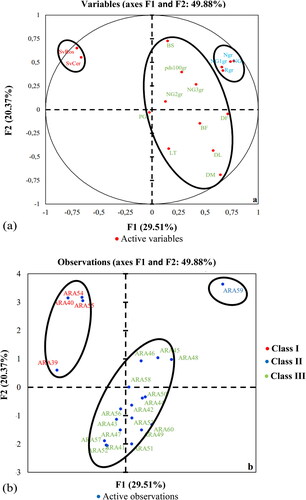
On the other hand, these variables are negatively correlated with the severity of late leaf spot and groundnut rosette, so the severity of these diseases limits the number of pods and seeds per plant, which harms the seed yield of peanut accessions affected by these diseases. In addition, these two diseases are positively linked, which increases parasite pressure on affected plants and limits their production potential. Axis 2 (F2), which accounts for 20.37% of total variability, is characterized by maturity delay (DM), which is negatively correlated with the severity of groundnut rosette and dry biomass (BS). When the time to maturity (DM) is late, the dry biomass and the severity of the disease increase.
Three groups of accessions were identified by combining the correlation circle () and the projection of individuals on the different axes (). Group I is explicitly characterized by its susceptibility to groundnut rosette (SvRos) and late leaf spot (SvCer) and includes accessions ARA40, ARA39, ARA54, and ARA55. Group II, made of only accession ARA59, is mainly characterized by the number of pods and seeds per plant and a good yield. Group III consists of accessions ARA45, ARA46; ARA48; ARA58; ARA44; ARA42; ARA50; ARA51; ARA53; ARA49; ARA60, ARA56; ARA43; ARA47; ARA57; ARA52, and ARA41. This group is characterized by a good length of the main stems (LT) of the plants, a high number of two-seeded pods (NG2gr) and three-seeded pods (NG3gr), a good weight of fresh pods (GF) and 100-seeded pods (pds100gr), a high dry biomass (BS) and fresh biomass (BF), and a late time to emergence (DL), flowering (DF) and ripening (DM).
Structuring and characterization of accessions studied in the second growing cycle
The parameters of vegetative growth, phenology, yield and components, and the severity of late leaf spot mark the second crop cycle. These parameters contributed to forming the first two axes (F1 and F2) at 72.63%. The first axis (F1) accounts for 58.89% of total variability. The variables that contribute to the formation of this axis are defined by the time to emergence (DL), flowering (DF) and maturity (DM), the length or height of the stem (LT) and the weight of fresh pods (PGF), number of pods per plant (NG/pl), number of one-seeded pods (NG1gr), number of seeds (Ngr), dry biomass (BS), seed yield (Rgr), severity of late leaf spot (SevCer). On this axis, the agromorphological parameters mentioned above are positively correlated, so an increase in these variables (length of the main stem, flowering time, weight of fresh pods, and dry biomass) simultaneously leads to an increase in seed yield. Conversely, seed yield fell when the pathogen of late leaf spot became more aggressive, leading to an increase in the severity rating ().
Figure 4. Correlation circle (a) and dispersion of accessions; (b) on the F1 and F2 axes in the second crop cycle.
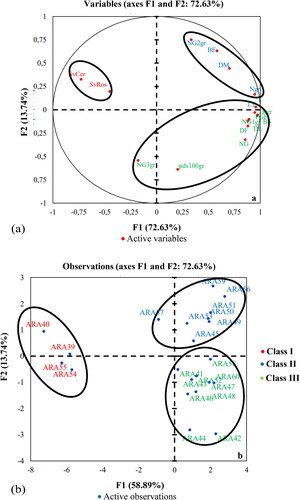
The second axis (F2) accounts for 13.72% of total diversity. It is characterized by dry biomass (BS). The combined observation of the correlation circle () and the distribution of individuals () on the axes revealed three (3) groups of accessions, as in the first crop cycle. The individuals making up group I are identical and have the same characteristics as in the first cropping cycle. Those in group II are characterized by a good seed yield (Rgr), large stems (LT), a longer time to emergence (DL) and flowering (DF), a higher dry biomass (BS), a higher number of one-seeded pods (NG1gr) and three-seeded pods (NG3gr) and a higher 100-seed weight (pds100). These are accessions ARA58, ARA41, ARA52, ARA43, ARA47, ARA46, ARA48, ARA60, ARA44, and ARA42. Group III, comprising accessions ARA59, ARA56, ARA51; ARA50, ARA49; ARA45; ARA53; and ARA57, is characterized by the number of two-seeded pods (NG2gr), the number of pods (NG), a higher fresh biomass per plant (BF) and late maturity (DM). However, these two sub-groups are resistant to groundnut rosette and susceptible to late leaf spot.
Groups of accessions studied in the first and second cycles using hierarchical ascending classification (HAC)
The hierarchical ascending classification (HAC) was used to estimate the degree of similarity between the groundnut accessions used. Accessions belonging to the same group have similar agromorphological and health parameters. Thus, the ascending hierarchical classification made it possible to discriminate between three distinct groups of individuals at each cropping cycle ().
Figure 5. Dendrograms showing the groups of accessions revealed with similarity levels of 50 in the first (a) and second; (b) growing cycle.
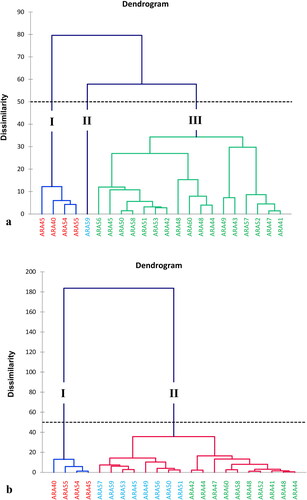
The dendrogram in confirmed the three groups obtained using PCA for the first crop cycle. Group I consist of accessions ARA39, ARA40, ARA54, and ARA55; group II consists solely of a single accession ARA59 and group III includes individuals ARA56, ARA45, ARA50, ARA58, ARA5, ARA53, ARA42, ARA48, ARA60, ARA46, ARA44, ARA49, ARA43, ARA57, ARA52, ARA57, and ARA41 (). In the second crop cycle, HAC revealed two (2) homogeneous groups within the peanut accessions, unlike PCA, which revealed three homogeneous groups. Group I had the same accessions (ARA40, ARA55, ARA54, and ARA39) as in the PCA. Group II was made up of accessions ARA57, ARA59, ARA53, ARA45, ARA49, ARA56, ARA50, ARA51, ARA42, ARA44, ARA47, ARA60, ARA58, ARA46, ARA52, ARA41, ARA48, and ARA43 (). However, HAC group II can be split into subgroups I and II. These two subgroups are identical to AMP groups II and III, respectively, with the same characteristics (). Subgroup 1: ARA57, ARA59, ARA53, ARA45, ARA49, ARA56, ARA50, and ARA51. Subgroup 2: ARA42, ARA44, ARA47, ARA60, ARA58, ARA46, ARA52, ARA41, ARA48, and ARA43.
Discussion
The agromorphological behavior of planted peanut accessions under late leaf spot and groundnut rosette stresses varied from the first to the second crop cycle on the same experimental site. This parameter variability concerns emergence, flowering and maturity times, plant length or height, yield, and its components. Generally speaking, the average time to reach the phenological stages recorded in the first cycle was shortened in the second growing cycle. The variability of these parameters between the same accessions at different growing cycles could be justified by the influence of biotic and/or abiotic factors covering the trial period. The average rainfall in the first crop cycle was less abundant (433.3 mm) than in the second cycle when it was 632 mm. A study by Tshilenge (2010) showed that climatic factors such as rainfall and temperature influence the expression of the disease (late leaf spot). This assertion is shared by Zinsou et al. (Citation2019), who showed that the incidence and mean severity varied from one agroecological zone to another. According to Koita et al. (Citation2017), plants affected by late leaf spot show a shortening of their vegetative cycle. This finding justifies the earlier emergence, flowering, and ripening times for accessions in the second crop cycle, especially as all peanut plants were severely attacked by late leaf spots, with mean severity ranging from 40 to 73.7% ().
It is therefore not surprising that the number of pods with one, two, and three seeds and seed yield are higher in the first growing cycle than in the second. Principal component analysis (PCA) shows that seed yield is strongly positively controlled by the number of pods per plant (NG), number of seeds (Ngr), number of pods to 1 seed (NG1gr), emergence time (DL), and flowering time (DF) and vegetative growth and development parameters (). These parameters were negatively correlated with late leaf spot and groundnut rosette. The occurrence of these diseases impairs these agromorphological parameters. These results are similar to those of Gadji et al. (Citation2021), who showed negative correlations (r = –0.62 and –0.69) between seed yield and the incidence of groundnut rosette and between seed yield and the severity of late leaf spot. Late leaf spot, caused by Cercosporidium personatum, is the most serious fungal disease of peanuts in Côte d’Ivoire (Blizoua Bi & Djan, Citation1995). It attacks all the aerial organs of the plant, particularly the leaves (Gadji et al., Citation2021). The disease causes multiple necrotic spots on the leaves, reducing the plant’s photosynthetic activity. Koita et al. (Citation2017) pointed out that pod filling requires good photosynthetic activity. This requirement is achieved thanks to the plant’s leaves, which intercept most light energy. A reduction in leaf area due to symptoms of late leaf spot and/or groundnut rosette leads to a reduction in the assimilation of CO2 required for synthesizing carbohydrates for better pod formation. The result is reduced pod yield, seed weight, and oil content (Wells et al., Citation1982). Therefore, the reduction in yield of the 22 peanut accessions studied is due to the virulence of fungal (late leaf spot) and viral (groundnut rosette) diseases in the second crop cycle between July and October. However, accessions ARA42 (1.59 t/ha), ARA60 (1.5 t/ha), ARA56 (1.49 t/ha), and ARA51 (1.42 t/ha) gave significant yields in the second cycle, unlike accessions ARA55 (0,131 t/ha), ARA40 (0.14 t/ha), ARA39 (0.231 t/ha), ARA54 (0.28 t/ha), ARA57 (0.713 t/ha) and ARA53 (0.996 t/ha) which gave the lowest seed yields. These results are similar to those obtained by Koita et al. (Citation2017). As mentioned above, these authors showed the negative impact of early and late leaf spots on peanut pod yield. On the other hand, the accessions ARA59 (5.05 and 1.399 t/ha) and ARA42 (2.72 and 1.594 t/ha) that yielded better in both cropping cycles were collected in Bouaké. Both accessions’ behavior suggests they are suited to their area of origin. Agroecological factors are thus influencing the agromorphological behavior of the accessions. Similarly, Zinsou et al. (Citation2019) assert that some varieties with adequate resistance in one area may become susceptible in another.
Concerning epidemiological parameters, principal component analysis (PCA) showed a strong positive correlation between late leaf spot and groundnut rosette irrespective of the crop cycle (). In the second crop cycle, Accessions ARA39, ARA55, ARA54, and ARA40 were the most affected by groundnut rosette, with mean severity ranging from 80 to 86.7%. Regarding late leaf spot, all accessions had the same attack level, with an average severity of 55.5%. The temperature in the second period was lower (25.17 °C) with higher rainfall (632 mm). On the other hand, in the first cycle, the temperature and rainfall were 27.34 °C and 433.3 mm, respectively. This would have favored a greater presence of groundnut rosette vectors and better development of C. personatum spores in the second crop cycle. Studies have shown that prolonged periods of high relative humidity and daily temperatures of 25 °C lead to better development of these two diseases in peanut plants (Zinsou et al., Citation2019; Gadji et al., Citation2021).
Furthermore, according to Ngoyi et al., (Citation2015), groundnut rosette and late leaf spot are seasonally dependent. This result would explain the variability in the susceptibility of accessions to groundnut rosette and late leaf spot during the growing season. This assertion is supported by Gadji et al. (Citation2021), who indicated that these diseases were more damaging in the second crop cycle than in the first. These claims contradict those of Hasan et al. (Citation1970), who argue that ecological factors, notably rainfall, even fine rainfall, can reduce Aphis populations, the vectors of peanut rosette. Thus, the severity of the peanut rosette should have been less marked during the second period.
The dispersion of the variables, which is visibly significant at the PCA level, attests to the existence of inter-accessional diversity. This diversity is manifested by three (3) distinct groups regardless of the crop cycle and would represent an important reservoir of interesting traits for other studies. Zinsou et al. (Citation2019) indicated that yield variability reflects the diversity of responses of varieties under growing conditions.
The HAC was used to estimate the degree of similarity between the peanut accessions used. Accessions belonging to the same group showed the same behavior for the agromorphological parameters studied. Accessions ARA39, ARA40, ARA54, and ARA55 from group I, then ARA42, ARA44, ARA47, ARA60, ARA58, ARA46, ARA52, ARA41, and ARA48 from group III are those that fluctuated less under the different climatic conditions. Accessions in group I and those that fluctuated from one group to another depending on the crop cycle were more susceptible to groundnut rosette and late leaf spot. On the other hand, those in group III were less susceptible to these diseases. These results are similar to those of Boni (Citation2017), who obtained three (3) groups using these same accessions over the two crop cycles. This author showed that group I accessions fluctuated the least and showed high susceptibility to groundnut rosette and late leaf spot for all crop cycles.
Conclusion
Our study highlighted the negative impact of late leaf spot and groundnut rosette on cultivated peanuts’ agromorphological and yield parameters. It showed that these diseases are more prevalent in the second crop cycle than in the first. This fact significantly reduced the production potential of peanuts grown on plantations in Central Côte d’Ivoire. The peanut accessions evaluated were susceptible to late leaf spots, and only four (ARA40, ARA54, ARA45, and ARA55) to groundnut rosette. The multivariate analyses distinguished three groups of peanut accessions based on their degree of similarity. Of these groups, one proved resistant or tolerant to peanut rosette, and two moderately resistant. Accessions ARA42 (1.59 t/ha), ARA60 (1.5 t/ha), ARA56 (1.49 t/ha), and ARA51 (1.42 t/ha), which were moderately resistant to groundnut rosette and susceptible to late leaf spot, obtained significant seed yields. On the other hand, ARA55, ARA40, ARA39, ARA54, ARA57, and ARA53, susceptible to these parasitic diseases, obtained the lowest seed yields. Accessions moderately susceptible to groundnut rosette and late leaf spots with good yields in the second crop cycle should be selected for further studies to refine the results obtained.
Authors’ contributions
This work was carried out in perfect collaboration between all the authors. MFDPN initiated the study, corrected the protocol, provided all the materials and documentation needed to carry out the work, and contributed to the drafting of the manuscript. AAGG developed the protocol, supervised the agricultural fieldwork and post-harvest activities, and drafted the manuscript. NDK carried out the statistical analyses of the data collected, contributed to the drafting of the manuscript, and participated in translating the manuscript into English. Author 4 (BUK) conducted the agricultural activities, collected the data, participated in the statistical analyses and contributed to the drafting of the manuscript. Author 5 (LF) contributed to the interpretation of the results and the drafting of the manuscript. Authors 6 (CLO), 7 (AFN), 8 (EOT), and 9 (KA) corrected the manuscript in French and English.
Public Interest Statement.docx
Download MS Word (16.4 KB)About the Authors.docx
Download MS Word (16.7 KB)Acknowledgments
The authors would like to thank the Centre National de Recherche Agronomique (CNRA) authorities, particularly those of the Research Station on Food Crops in Bouaké, for having made plots available to us to set up the experiment. We would also like to thank Mr. KANATE Issouf, Senior Technician at the CNRA, and Mr. KOUASSI Kouakou, Specialist Worker at the CNRA, for setting up the peanut trial.
Disclosure statement
The authors have no known competing interests.
Data availability statement
The information presented in this study is available on request from the corresponding author.
Correction Statement
This article has been corrected with minor changes. These changes do not impact the academic content of the article.
Additional information
Funding
Notes on contributors

Kouabenan Abo
This manuscript is based on the results of research carried out by the Vegetable and Protein Crops Program of the Centre National de Recherche Agronomique (CNRA) in Côte d’Ivoire. This program is made up of researchers of various specialities holding a PhD from the Université Félix Houphouët-Boigny de Cocody and the Université Nangui Abrogoua (Côte d’Ivoire). There are two researchers in Plant Genetics and Breeding, two researchers in Agro-Physiology, one researcher in Plant Pathology and Eco-Health and one researcher in Entomology. The aim of the research program is to contribute to a sustainable increase in market garden and protein crop production in Côte d’Ivoire. To achieve this, the research team is working to improve the productivity and health quality of vegetable and protein crops, including peanuts. The research team works in collaboration with several national and international agricultural research institutions and receives funding from national and international organisations via the Consultative Group on International Agricultural Research (CGIAR).
Photo of the authors and technicians
The researchers are white coats and technicians in blue coats.
References
- Blizoua Bi, P. Z., & Djan, N. V. (1995). Pertes de rendements chez l’arachide dues à un déficit hydrique en relation avec la cercosporiose tardive (Cercosporidium personatum) en Côte d’Ivoire. Cahiers Agricultures, 4, 1–12.
- Boni, A. (2017). Incidence de la cercosporiose et de la rosette sur quelques caractères de production de vingt et une accessions d’arachide (Arachis hypogaea L.) cultivées dans la région de Bouaké (Centre de la Côte d’Ivoire). Mémoire de Master, Université Felix Houphouët Boigny, Abidjan, Côte d’Ivoire (p. 57).
- Christie, M. E., Kyamureku, P., Kaaya, A., & Devenport, A. (2015). Farmers, peanuts, and aflatoxins in Uganda: A gendered approach. Development in Practice, 25(1), 4–18. https://doi.org/10.1080/09614524.2015.983459
- Coulibaly, N. D., Ossey, C. L., Gadji, A. G., De Paul N’Gbesso, M. F., Fondio, L., & Soro, O. T. (2021). Etude De L’arrière Effet Des Légumineuses Alimentaires Sur La Productivité Des Légumes : Cas De La Tomate (Solanum Lycopersicum), Cultivée Dans La Localité De Bouaké Au Centre De La Côte d’Ivoire. European Scientific Journal, ESJ, 17(21), 125 https://doi.org/10.19044/esj.2021.v17n21p125
- FAO. (2014). Food and Agriculture Organization of the United Nations, Statistics Division, http://www.fao.org/faostat/fr/#data/QC. Accessed on 27 October 2019.
- FAOSTAT. (2020). Agricultural production, crop primary database, Food and Agricultural Organization of the United Nations, http://faostat.fao.org/faostat. Accessed on 14 July 2021.
- Gadji, A. A. G., Kouame, K. G., Coulibaly, N. D., Ossey, C. L., N’Gbesso, M. F. P., & Fondio, L. (2021). Impact de la cercosporiose et de la rosette sur les performances agronomiques de 12 accessions d’arachide (Arachis hypogaea L.) au Centre de la Côte d’Ivoire. Afrique Science, 19(5), 51–65.
- Guchi, E., Ayalew, A., Dejene, M., Ketema, M., Asalf, B., & Fininsa, C. (2014). Occurrence of Aspergillus species in groundnut (Arachis hypogaea L.) along the value chain in different agroecological zones of Eastern Ethiopia. Journal Applied Environmental Microbiology, 2, 309–317.
- Hasan, M. R., Ahmad, M., Rahman, M. H., & Haque, M. A. (1970). Aphid incidence and its correlation with different environmental factors. Journal of the Bangladesh Agricultural University, 7(1), 15–18. https://doi.org/10.3329/jbau.v7i1.4791
- James, W. C. (1974). Assessment of plant diseases and losses. Annual Review of Phytopathology, 12(1), 27–48. https://doi.org/10.1146/annurev.py.12.090174.000331
- Koita, K., M’Bi Zagre, B., & Sankara, P. (2017). Aqueous plant extracts for control of groundnut leaf spot in Burkina Faso. African Crop Science Journal, 25(3), 311–319. https://doi.org/10.4314/acsj.v25i3.5
- Manizan, L., Akaki, D., Piro-Metayer, I., Montet, D., Brabet, C., & Koffi-Nevry, R. (2018). Évaluation des pratiques culturales de l’arachide favorisant la contamination par les aflatoxines dans trois régions de Côte d’Ivoire. International Journal of Biological and Chemical Sciences, 12(4), 1590–1600. https://doi.org/10.4314/ijbcs.v12i4.7
- Marloire, M. (2019). Arachide, Encyclopædia Universalis [en ligne]. Available at: http://www.universalis.fr/encyclopedie/arachide/. Accessed: 29 December 2020.
- N’Gbesso, M. F. D. P., Fondio, L., Coulibaly, N. D., Djidji, A. H., & Dibi, K. E. B. (2020). Bien cultiver l’arachide en Côte d’Ivoire. Fiche Plante Protéagineuse, 2, 4.
- Ngoyi, F., Mudibu, W., Kabangu, J., Tshiombe, M. V., & Masiala, M. G. (2015). Identification of adapted varieties of groundnuts (Arachis hypogaea L.) in SEKE BANZA area, Democratic Republic of the Congo (DRC). Int. J. Biol. Chem. Sci, 9(2), 652–663.
- Okello, D. K., Biruma, M., & Deom, C. M. (2010). Overview of groundnuts research in Uganda: past, present and future. African Journal Biotechnology, 9(39), 6448–6459.
- Olorunju, P. E. (1991). Disease Reactions and Yield Performance of Peanut Genotypes Grown Under Groundnut Rosette and Rosette-Free Field Environments. Plant Disease, 75(12), 1269. https://doi.org/10.1094/PD-75-1269
- Reddy, E. C., Sudhakar, C., & Reddy, N. P. (2011). Aflatoxin contamination in groundnut induced by Aspergillus flavus type fungi: A critical review. http://imsear.li.mahidol.ac.th/handle/123 456789/161664.
- Schilling, R., Dimanche, P., Crambade, P., & Gautreau, J. (1996). ‘‘L’arachide en Afrique Tropicale’’, Ed. Maisonneuve et Larose CTA, Paris, 25–35.
- Van Lenteren, J. C., Bolckmans, K., Köhl, J., Ravensberg, W. J., & Urbaneja, A. (2018). Biological control using invertebrates and microorganisms: plenty of new opportunities. BioControl, 63(1), 39–59. https://doi.org/10.1007/s10526-017-9801-4
- Wells, R., Schulze, L. L., Ashley, D. A., Boerma, H. R., & Brown, R. H. (1982). Cultivar differences in canopy apparent photosynthesis and the relationship to seed yield in soybeans. Crop Science, 22(4), 886–890. https://doi.org/10.2135/cropsci1982.0011183X002200040044x
- Zinsou, V. A., Garba Say, L., Fanou, A., Sekloka, E., Koussinou, C., Djenontin, A. J., Aweha, A., Dramane, M., Korogone, C., & Gagnon, F. (2019). Importance des cercosporioses de l’arachide et sélection des variétés pour la résistance aux maladies au Bénin. Tropicultura, 37(4), 4–22.