
Abstract
Osteoblasts (OBs), which are a crucial type of bone cells, derive from bone marrow mesenchymal stem cells (MSCs). Accumulating evidence suggests inflammatory cytokines can inhibit the differentiation and proliferation of OBs, as well as interfere with their ability to synthesize bone matrix, under inflammatory conditions. NLRP3 inflammasome is closely associated with cellular pyroptosis, which can lead to excessive release of pro-inflammatory cytokines, causing tissue damage and inflammatory responses, however, the comprehensive roles of NLRP3 inflammasome in OBs and their differentiation have not been fully elucidated, making targeting NLRP3 inflammasome approaches to treat diseases related to OBs uncertain. In this review, we provide a summary of NLRP3 inflammasome activation and its impact on OBs. We highlight the significant roles of NLRP3 inflammasome in regulating OBs differentiation and function. Furthermore, current available strategies to affect OBs function and osteogenic differentiation targeting NLRP3 inflammasome are listed and analyzed. Finally, through the prospective discussion, we seek to provide novel insights into the crucial role of NLRP3 inflammasome in diseases related to OBs and offer valuable information for devising treatment strategies.
Introduction
Skeletal development and bone remodeling rely on a delicate balance between the bone formation process executed by osteoblasts (OBs) and the bone resorption process carried out by osteoclasts (OCs),Citation1 OCs initiates the resorption of existing bone, thus triggering a subsequent process of bone formation by OBs.Citation2 During bone remodeling, bone marrow mesenchymal stem cells (MSCs) undergo differentiation into OBs, which subsequently mature into osteocytes.Citation3 Importantly, many diseases, such as osteoporosis (OP),Citation4,Citation5 periodontitis,Citation6 osteoarthritis (OA),Citation7,Citation8 are characterized by the disruption of bone remodeling processes, furthermore, calcification, a risk factor for the occurrence of multiple diseases such as atherosclerosis and nephrocalcinosis,Citation9,Citation10 is highly related to osteogenic differentiation, in calcified blood vessels or tissues, many cells become activated and undergo a process of trans-differentiation into other cell types, such as OBs, which express bone markers and matrix proteins including osteopontin (OPN) and runt-related transcription factor 2 (RUNX2), which ultimately lead to calcification.Citation11,Citation12 Therefore, it is essential to further elucidate mechanisms that affect OBs function and differentiation to facilitate the development of novel therapeutic strategies for these diseases.
Figure 1 A schematic model depicting the activation process of NLRP3 inflammasome and the impact on OBs, osteogenic differentiation and osteogenic microenvironment. Various intracellular and extracellular stimuli activate NLRP3 inflammasome, leading to the release of inflammatory factors and alterations in the expression of markers associated with osteogenic differentiation. Consequently, this impedes the functionality and osteogenic differentiation capacity of OBs and their precursor cells, MSCs.
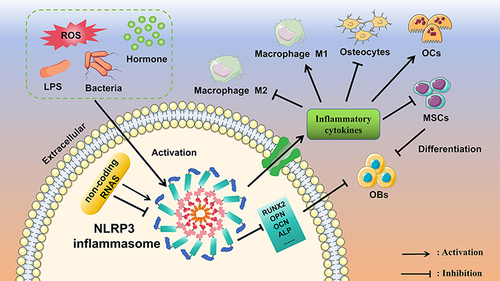
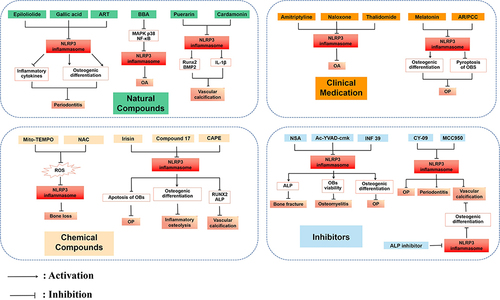
Figure 2 Potential therapeutic strategies by inhibiting NLRP3 inflammasome in OBs-related diseases.
Pyroptosis, an inflammatory and programmed mode of cell death (PCD), is composed of “pyro” and “ptosis”. “Pyro” means fire, indicating the properties of inflammation, and “ptosis” means falling, which is consistent with other forms of programmed cell death.Citation13 This process is inherently dependent on inflammation and can be triggered by various pathological stimuli, such as stroke, heart attack, or cancer. It plays a crucial role in controlling microbial infections.Citation14 Importantly, as an intracellular multiprotein complex, the NLRP3 (NOD-, LRR- and pyrin domain-containing protein 3) inflammasome plays a crucial role during the process of pyroptosis, the formation and activation of the NLRP3 inflammasome leads to release of the pro-inflammatory cytokines interleukin (IL) −1β and IL-18 as well as to the pyroptotic cell death.Citation15 Notably, recent studies have revealed that the activation of NLRP3 inflammasome can have a significant impact on the function and differentiation of OBs. This activation has been linked to increased bone resorption, impaired mineralization, and decreased bone formation.Citation16,Citation17 Furthermore, studies have demonstrated that drugs, natural compounds as well as inhibitors can promote OBs differentiation by decreasing the NLRP3 inflammasome.Citation18–21 However, the precise roles of NLRP3 inflammasome in OBs and their differentiation have not been fully established. This lack of consensus has hindered the development of novel insights into the therapeutic application of targeting NLRP3 inflammasome in diseases associated with OBs and their differentiation.
In this review, we provide a concise overview of the activation of NLRP3 inflammasome and briefly introduce the effects of NLRP3 inflammasome activation on OBs and osteogenic microenvironment, which emphasizes the crucial roles of NLRP3 inflammasome on differentiation and function of OBs. Additionally, we present and examine the existing approaches that can impact the function of OBs and their differentiation by targeting the NLRP3 inflammasome. Moreover, our prospective discussion aims to provide fresh perspectives on the significant role of the NLRP3 inflammasome in OBs-related diseases and furnish valuable information for the development of treatment strategies.
The NLRP3 Inflammasome Pathway
The concept of the inflammasome was first proposed in 2002, it is a multi-protein complex discovered in the innate immune system.Citation22 The basic structure of inflammasome is composed of NOD-like receptor family (NLRs) proteins as sensor proteins, apoptosis-associated speckle-like protein containing a caspase recruitment domain (ASC) as an adaptor protein, and caspase-1 as an effector protein. The assembly of the inflammasome depends on identifying pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) by pattern recognition receptors (PRRs).Citation23 Due to the relatively fixed structure of ASC and caspase-1, while the types of sensor proteins are diverse, the inflammasomes are classified according to the sensor proteins. To date, NLRP1, NLRP3, and NLRC4 from the NLRs, absent in melanoma 2 (AIM2) from the AIM2-like receptor family (ALRs) and pyrin from the tripartite motif-containing protein family are the best-characterized sensors reported to form inflammasome complexes.Citation24
NLRP1 was the initial inflammasome-forming sensor discovered at the turn of the century and is now acknowledged as the primary inflammasome sensor protein in human keratinocytes.Citation25,Citation26 NLRC4 was initially characterized as a pro-apoptotic protein. Over the last decade, research has highlighted its crucial role in combating microbial infections and maintaining homeostasis, although this function can vary depending on the type of pathogen and the level of infection. NLRC4 also exerts immunomodulatory effects during bacterial infections. Several gram-negative bacteria, such as Legionella pneumophila, Pseudomonas aeruginosa, Salmonella typhimurium and Shigella flexneri, activate caspase-1 and induce pyroptosis through the NLRC4 inflammasome.Citation27,Citation28 AIM2, a cytoplasmic sensor that detects double-stranded DNA, is involved in various physiological processes, including development, infectious diseases, inflammatory conditions, and cancer. AIM2-mediated release of effector cytokines and induction of pyroptosis play crucial roles in defending the host against microbial infections.Citation29,Citation30 Pyrin, encoded by the MEFV gene, is primarily found in innate immune cells within the cytoplasm. It consists of five domains, each with a specific function in modulating cytokine secretion, cellular death pathways, and cytoskeletal signaling. These domains interact with various proteins to regulate these cellular processes.Citation31,Citation32 Pyrin activation relies heavily on molecular events that disrupt the function of small Ras Homolog Family Member A (RhoA) GTPases, primarily through bacterial toxins or bacterial effectors. This process alters cellular homeostasis, ultimately triggering pyrin activation.Citation33,Citation34 Excessive pyrin activation is associated with inflammatory disorders such as Mediterranean Fever and pyrin-associated autoinflammation with neutrophilic dermatosis.Citation35
The most studied member of inflammasome complexes is NLRP3. NLRP3 is a sensor protein, which is composed of three domains: a pyrin domain (PYD) at the amino-terminal, a NACHT domain with ATPase activity in the nucleotide-binding region, and a leucine-rich repeat (LRR) domain at the carboxy-terminal.Citation36 NLRP3 inflammasome is a prominent inflammatory complex that includes NLRP3, ASC, and caspase-1. Upon activation of this complex, it initiates the activation of caspase-1, which subsequently cleaves pro-IL-1β and pro-IL-18, leading to the production of mature forms of these inflammatory cytokines, namely IL-1β and IL-18. This activation cascade triggers and amplifies inflammatory responses within the body.Citation37,Citation38 Simultaneously, in response to microbial infection and cellular damage, it cleaves Gasdermin D (GSDMD) into its active form, and features early plasma membrane rupture, thereby releasing the soluble intracellular fraction that initiates pyroptosis.Citation39–41
As of now, research has demonstrated that NLRP3 inflammasomes can be activated via two distinct signaling pathways: canonical NLRP3 inflammasome activation and noncanonical NLRP3 inflammasome activation. The former is currently believed to involve three stages: priming, activation, and post-translational modification (PTM)-interacting components. During the priming step, the initial signal such as lipopolysaccharide (LPS) triggers the activation of toll-like receptors (TLRs) and nuclear factor-kappa B (NF-κB) and subsequently produces NLRP3 and pro- IL-1β. During the following activation step, multiple stimuli, such as ATP and nigericin trigger the assembly of NLRP3 inflammasome.Citation15,Citation42 However, certain studies suggested that under certain circumstances, particular triggers could directly activate the inflammasome without requiring a preceding priming step. For instance, the activation of NLRP3 by nigericin triggers the processing and release of constitutively expressed IL-18, even in the absence of priming.Citation43,Citation44 PTMs of NLRP3 induce inflammasome activation and determine the inflammation intensity by affecting the protein stability, ATPase activity, subcellular localization, and oligomerization of NLRP3 as well as the association between NLRP3 and other inflammasome components, several PTMs of NLRP3 were reported, such as ubiquitylation, deubiquitination, phosphorylation, sumoylation and palmitoylation.Citation45 The latest research indicates acetylation is critical for the activation of NLRP3 inflammasome in response to diverse stimuli.Citation46 Multiple proteins participate in a plethora of PTMs of NLRP3 inflammasome components, modifying their protein functions, activities, and/or intracellular localization.Citation47 The latter results from the sensing of intracellular LPS through a mechanism that depends on TLR4 and type I interferons, as well as guanylate-binding proteins, leading to activation of caspase-11. Caspase-11 facilitates GSDMD activation and split, then mediates pyroptosis. Additional comprehensive insights into the mechanism of NLRP3 inflammasome activation can be found in preceding reviews.Citation48–50 Notably, the mitotic serine/threonine kinase NEK7 has emerged as a crucial factor in NLRP3 activation, as it directly interacts with NLRP3.Citation51 Moreover, the NLRP3 inflammasome is extensively researched and widely acknowledged as being activated by numerous viral families. Studies have demonstrated its activation by various RNA viruses across different families.Citation52
The aberrant activation of the NLRP3 inflammasome contributes to various inflammatory diseases including chronic colitis,Citation53 endothelial dysfunction,Citation54 periodontitis,Citation55 OP Citation56 and others. Therefore, studying the activation mechanisms of NLRP3 inflammasome in related diseases and identifying and evaluating drugs that can target NLRP3 inflammasome hold promise for the treatment of these conditions. However, due to the intricate interplay and overlapping nature of numerous upstream signals involved in NLRP3 inflammasome activation, there remains a lack of consensus regarding its precise mechanism of activation.
NLRP3 Inflammasome on OBs
OBs, which are responsible for bone formation, originate from MSCs that commit to osteoprogenitor lineages. This process involves the sequential activation of transcription factors and ultimately leads to the terminal differentiation of OBs into osteocytes.Citation57,Citation58 OBs play a pivotal role in bone tissue formation. They are responsible for producing and releasing a range of extracellular matrix proteins, including alkaline phosphatase (ALP). Several OBs come together to form osteons, where calcium is deposited as hydroxyapatite along with type I collagen, contributing to the structural integrity of the skeleton.Citation59,Citation60 Function of OBs is influenced by a myriad of factors, including oxidative stress, hormonal regulation, inflammatory cytokines, growth factors and cellular signals, exercise and mechanical stimulation, as well as senescence and epigenetic factors.Citation59,Citation61–63 Noteworthy, reports have begun to emerge regarding the impact of NLRP3 inflammasome activation induced by these factors on OBs function.
It is well-established that inflammatory responses can be initiated in the presence of oxidative stress and reactive oxygen species (ROS),Citation64,Citation65 leading to the activation of the NLRP3 inflammasome and subsequent OBs loss.Citation66 Another study similarly found that oxidative stress caused by LPS induces NLRP3 inflammasome-mediated pyroptosis of OBs, resulting in decreased cell migration as well as osteogenic dysfunction.Citation67 IL-17 is a pro-inflammatory cytokine that can activate NLRP3/ASC/caspase-1-dependent IL-1β processing and pyroptosis in OBs, leading to the release of cytoplasmic contents, including RANKL and IL-1β. This process can accelerate inflammatory bone destruction.Citation68 Estrogen plays a vital role as the primary hormonal regulator of bone metabolism in both women and men, its direct effects on OBs contribute to the maintenance of bone formation.Citation69 Research studies have shown that a deficiency of estrogen can lead to the activation of the NLRP3 inflammasome.Citation70 Studies have demonstrated that inhibiting the NLRP3-dependent NF-κB and MAPKs signaling pathways can alleviate bone loss caused by ovariectomy (OVX) in mice.Citation70 Likewise, in diabetic and OVX rats, there was an increased presence of ANXA1/FPR2-positive OBs accompanied by a higher number of OBs expressing COX2, NLRP3, and IL-1β. These findings suggest that the ANXA1/FPR2 pathway may serve as a regulatory mechanism to counterbalance the heightened activation of COX2, NLRP3, and IL-1β in bone cells during bone remodeling, particularly under conditions of estrogen deficiency and diabetes mellitus. This highlights a potential role for the ANXA1/FPR2 pathway in fine-tuning and regulating the inflammatory response in bone cells.Citation71 Moreover, all-trans retinoic acid (ATRA), a main derivative of vitamin A, could disrupt the osteogenesis and mineralization of periodontal ligament stem cells by promoting IL-1β expression via activating NF-κB signaling and NLRP3 inflammasome.Citation72
In addition, multipathogenic bacteria play an important role in activating the NLRP3 inflammasome, which can significantly affect OBs function. McCall et al observed that exposure of OBs to Salmonella led to a notable increase in NLRP3 protein expression. Importantly, when OBs were transfected with siRNA targeting NLRP3, the bacterially induced decrease in NF-κB activity was notably reduced, indicating that the ability of Salmonella to suppress the activity of NF-κB, an anti-apoptotic factor, is largely mediated by the presence of NLRP3.Citation73 Infection of OBs with Aggregatibacter actinomycetemcomitans (A. actinomycetemcomitans) resulted in the activation of NLRP3 inflammasomes, leading to the secretion of mature IL-1β and IL-18. Knockdown of NLRP3 expression using specific siRNA reduced the level of apoptosis observed in A. actinomycetemcomitans-infected OBs. These findings also underscore that bacterium may promote apoptosis of OBs at least partially through activation of the NLRP3 inflammasome.Citation74 Similarly, the presence of Enterococcus faecalis caused the induction of apoptosis and pyroptosis in OBs through the activation of the NLRP3 inflammasome, which, in turn, led to a delay in the reconstruction of periapical lesions.Citation75 Porphyromonas gingivalis, an oral bacterium, activated the double-stranded RNA-dependent kinase (PKR), which in turn increased NLRP3 expression by activating NF-κB in OBs, suggesting that PKR, along with NLRP3 affects inflammation in OBs during periodontal diseases.Citation76 Beyond that, upon stimulation by Staphylococcus aureus, the NLRP3 inflammatory sensor in OBs becomes activated, triggering caspase-1 activation. Caspase-1 cleaves GSDMD, resulting in the formation of membrane pores. These pores contribute to cellular swelling, eventual cell lysis, and the release of inflammatory factors such as IL-1β and IL-18 within OBs.Citation77
Apart from that, non-coding RNA can also influence the expression of NLRP3 inflammasome, thus impacting the function of OBs. It was clearly shown that NEAT1, an important long non-coding RNA (lncRNA), inhibited the NLRP3-mediated inflammatory process and hindered the function of OBs. This was achieved by enhancing autophagy and reducing the levels of caspase-1 and IL-1β expression.Citation78 Interestingly, another lncRNA, TCONS_00072128 significantly activated inflammation pathways including NLRP3 signaling and NF-κB signaling in MSCs.Citation79 Hence, additional research is imperative to fully elucidate the influence of lncRNA on OBs regarding the induction of NLRP3 inflammasome activation. MicroRNAs (miRNAs) also broadly regulate normal biological functions of OBs and the progression of bone remodeling through NLRP3 inflammasome, for example, miRNA-150-5p aggravated pyroptosis-dependent skeletal loss and the inflammatory response.Citation19 And miRNA-30a effectively suppressed NLRP3 inflammasome activation, reduced joint inflammation, and attenuated bone damage in TNFTG mice, the population of OBs and the ALP-positive surface were significantly decreased in NLRP3KO/TNFTG mice compared with TNFTG mice.Citation80 Unexpectedly, by using NLRP3 knock-out mice, Detzen et al demonstrated NLRP3 played a role in bone formation by controlling the maturation of hypertrophic chondrocytes and the activity of OBs, OBs lacking NLRP3 expression exhibited impaired mineralization, along with a decrease in the expression of bone sialoprotein.Citation17 However, the prevailing consensus among studies is that the NLRP3 inflammasome exerts a detrimental effect on OBs. Activation of the NLRP3 inflammasome has been shown to promote adipogenic differentiation while suppressing osteogenesis.Citation81–84
NLRP3 Inflammasome on Osteogenic Differentiation
Osteogenic differentiation, also referred to as osteoblastogenesis, is regulated by various transcription factors. These include RUNX2, osterix, activating transcription factor 4, special AT-rich sequence-binding protein 2, and activator protein-1.Citation62,Citation85 It is a key process in bone formation, and its dysfunction leads to bone metabolism-related diseases.Citation86 A growing body of research suggests that the inflammatory microenvironment influences osteogenic differentiation, thereby impairing osteoblastic bone formation and contributing to decreased bone quantity and quality,Citation87 although current studies are primarily performed in vitro to analyze the effect of TNF-α and IL-1β on osteogenic differentiation.Citation88,Citation89 Nevertheless, the role and mechanism of NLRP3 inflammasome activation is also believed to participate in the process of osteogenic differentiation.
It has been reported that NLRP3 protein expression was maintained throughout the differentiation of OBs.Citation90 Of note, previous studies have demonstrated the differentiation capability of MSCs into OBs or adipocytes was determined after NLRP3 inflammasome activation, the osteogenic differentiation of MSCs was markedly decreased following activation of the NLRP3 inflammasome,Citation91 and osteogenic differentiation of mandibular MSCs was increased both in vitro and in vivo after specifically knockdown NLRP3 or using MCC950, a potent highly specific small molecule inhibitor of NLRP3.Citation83 Similarly, Liu et al revealed that MCC950 restored the expression of osteogenic differentiation-related proteins such as COL1, RUNX2 and ALP in OBs.Citation67 In contrast, CY-09, another inhibitor of NLRP3, effectively reduced the increased expression of NLRP3, caspase-1, and IL-1β in the NLRP3 inflammasome pathway. However, it attenuated the upregulation of osteogenic calcification markers such as RUNX2, SPARC, and BMP2 in stenotic valves. Moreover, CY-09 inhibited the differentiation of valvular interstitial cells (VICs) into OBs-like cells.Citation20 In a study on OP, it was found that NLRP3 knockout had an inhibitory effect on the inflammatory response, leading to enhanced osteogenic differentiation of OBs) in mice with OVX.Citation92 In parallel, in other bone metabolism-related diseases such as rheumatoid arthritis and periodontitis, the activation of NLRP3 inflammasome is also believed to be negatively correlated with osteogenic differentiation, inhibiting NLRP3 inflammasome could greatly rescue osteogenic differentiation.Citation16,Citation93,Citation94
In addition, certain important biological molecules also play a crucial role in the activation of the NLRP3 inflammasome and have an impact on osteogenic differentiation. The upregulation of elongator complex protein 2 (ELP2), a crucial protein in the JAK-STAT3 pathway, occurs during the pyroptosis of MC3T3-E1 cells, whose upregulation inhibits osteogenic differentiation by activating NLRP3.Citation95 Osteoblastic-specific FoxO1 knockout enhanced the expression of NLRP3 inflammasome signaling, leading to an amelioration in alveolar bone loss and manifesting as enhanced osteogenic potential.Citation16 Additionally, SOCS1, a suppressor of NLRP3, showed negative regulation of osteogenic differentiation in calcific aortic valve disease,Citation96 whereas VEGF and CGRP exerted a positive effect on the proliferation and differentiation of OBs. They were found to reduce apoptosis, stimulate mineralization, and enhance ALP activity by regulating the NLRP3 inflammasome pathway.Citation94,Citation97
NLRP3 Inflammasome on Osteogenic Microenvironment
In the past few years, there has been increasing recognition of the vital role that the cellular microenvironment plays in maintaining the balance, repair, and regeneration of skeletal tissues.Citation98,Citation99 The osteogenic microenvironment is a crucial element of this system, consisting of cellular components like OBs, OCs, and osteocytes, along with non-cellular components such as the extracellular matrix.Citation100 Moreover, as bone immunology advances, increasing evidence suggests that bone regeneration involves intricate interactions between various systems, notably the osteogenic microenvironment and immune systems, rather than just bone formation and resorption.Citation101 Immune cells, such as macrophages, play dynamic roles in both normal bone physiology and pathological conditions by secreting cytokines that exert significant regulatory influence on the processes of osteoclastogenesis and osteogenesis.Citation102 Therefore, fully understanding the impact of the NLRP3 inflammasome on the osteogenic microenvironment can provide significant insights for the development of therapeutic strategies for inflammation-related bone disorders.
Osteocytes are cells living within the bone-mineralized matrix, which regulate both the OBs and OCs activities during bone remodeling. Blocking NLRP3 inflammasome activation or decreasing ROS production prevented the pyroptotic death of osteocytes, resulting in reduced osteoclastogenesis and periprosthetic osteolysis.Citation103,Citation104 OCs originate from myeloid progenitor or osteal macrophages and are responsible for bone resorption.Citation105 Multiple studies have indicated that the NLRP3 inflammasome can promote the maturation and differentiation of OCs, causing exceeding bone resorption.Citation106–109 It means that NLRP3 inflammasome-mediated OCs enhancement acts as a major driver of inflammatory bone loss. Macrophages, integral to bone immunity, exhibit distinct polarization into either pro-inflammatory M1 or anti-inflammatory M2 phenotypes in response to environmental cues, thereby intricately regulating bone metabolism and contributing to diverse functions in bone homeostasis.Citation110,Citation111 M1 macrophages serve dual roles: they not only serve as precursors for OCs and undergo differentiation into mature OCs but also secrete pro-inflammatory cytokines to enhance bone resorption. Conversely, M2 macrophages secrete osteogenic factors, fostering the differentiation and mineralization of OBs precursors and MSCs, consequently promoting bone formation.Citation112 Zhu et al found that STAT3 triggered Macrophage NLRP3 inflammasome activation, which released IL-1β to favorably enhance RANKL-induced osteoclastogenesis and bone-resorptive function.Citation113 In parallel, inhibiting NLRP3 expression could promote bone regeneration and facilitate angiogenesis by inhibiting M1 macrophage polarization or increasing the infiltration of M2 polarized macrophages.Citation114,Citation115 Similarly, in rheumatoid arthritis and periodontitis, downregulating the expression of NLRP3 and caspase-1, thereby preventing inflammatory cell death resulting from the release of IL-1β and IL-18 contributes to inhibition of M1 polarization and pyroptosis of macrophages.Citation116,Citation117 Previous studies have reported that M1 macrophages could secrete pro-inflammatory cytokines, which promotes an OBs-like phenotype in VICs.Citation118 Thus, inhibition of NLRP3 activity not only prevented the polarization of macrophages toward the M1 phenotype but also reduced the levels of pro-inflammatory factors such as IL-6 and TNF-α. This intervention led to improved aortic valve function and decreased deposition of calcification in the valve.Citation20 However, it should be noted that in inflammatory contexts, the traditional M1/M2 classification fails to fully capture the intricate functions of macrophages during osteoclastogenesis and osteogenesis, highlighting the need for a more comprehensive understanding.
Potential Therapeutic Strategies Targeting NLRP3 Inflammasome in OBs-Related Diseases
Given the importance of NLRP3 inflammasome in OBs, osteogenic differentiation and osteogenic microenvironment (), various therapeutic strategies targeting NLRP3 inflammasome have been applied in vivo and in vitro to treat OBs-related diseases (), including natural compounds such as artesunate (ART), puerarin, epiloliolide, chemical compounds such as N-acetylcysteine (NAC), compound 17, clinical medication such as amitriptyline, melatonin, and IL-1 inhibitors, and protein inhibitors such as necrosulfonamide (NSA), Ac-YVAD-cmk, MC950.
Table 1 Therapeutic Agents Targeting NLRP3 Inflammasome in Diseases Associated with OBs and Osteogenic Differentiation
Natural Compounds
ART, which is derived from Artemisia annua L., has been found to possess anti-inflammatory and immunomodulatory properties, making it valuable in the treatment of various immune and chronic diseases.Citation132 Furthermore, ART has shown dose-dependent effectiveness in improving ligature-induced periodontitis by enhancing osteogenic differentiation, subsequent RNA-seq analysis has indicated that ART effectively decreases the expression of genes associated with the NLRP3 inflammasome.Citation119 Vascular calcification increases vascular stiffness, lowers organ perfusion, and augments cardiac afterload, thus resulting in left ventricular hypertrophy, diastolic dysfunction and heart failure. Vascular calcification is closely related to osteogenic differentiation.Citation133 Puerarin is a major bioactive ingredient derived from the root of Pueraria lobata (Willd.) Ohwi, it could decrease the expression of osteogenic differentiation markers RUNX2 and BMP2 by targeting NLRP3/ caspase1/IL-1β in mouse vascular smooth muscle cells, which shows great potential to treat vascular calcification clinically.Citation120 That which has a similar effect is Cardamonin, it exhibited anti-inflammatory and anti-calcification effects in human VICs by suppressing the activation of the NLRP3 inflammasome and downregulating IL-1β expression.Citation121 Furthermore, β boswellic acid (BBA) binds to the innate immune receptor TLR4 complex and inhibits OA-associated inflammatory and catabolic processes in OBs. It inhibits TLR4 and IL1R signaling and downregulates MAPK p38/NF-κB, NLRP3 inflammasome, and other OA-associated pathways.Citation122 Epiloliolide has been verified to enhance the growth, movement, and ability of human periodontal ligament cells to transform into OBs while reducing the production of pro-inflammatory mediators and cytokines including iNOS, COX-2, TNF-α, IL-6, and IL-1β by modulating the activity of NLRP3.Citation123 In the same way, Gallic acid could eliminate the inflammatory cytokines (such as IL-6 and IL-1β) and targets of the inflammasome (including Caspase-1 and NLRP3) induced by Pg-LPS in human ligament periodontal cells. Additionally, it was observed to enhance ALP activity and mineralization, which consequently facilitated the process of OBs differentiation.Citation21
Chemical Compounds
Bisphenol A (BPA), a widely-present chemical that disrupts the endocrine system, was found to reduce the viability of MLO-Y4 cells and induce apoptosis in a dose-dependent manner. This effect was attributed to the activation of the ROS/NLRP3/Caspase-1 pathway. Interestingly, the activation of the NLPR3 inflammasome and subsequent pyroptotic death caused by BPA was effectively inhibited by the ROS scavenger NAC or the mitochondrial antioxidant Mito-TEMPO.Citation104 Irisin, a myokine that acts like a hormone, can increase the expression of Nrf2, suppress the activation of the NLRP3 inflammasome, and decrease the levels of inflammatory factors. This mechanism ultimately led to the inhibition of OBs apoptosis in postmenopausal OP rats and contributed to a reduction in the incidence of postmenopausal OP.Citation82 Besides, a small molecule named compound 17, promoted the mineralization, ALP activity as well as the mRNA expression of OBs differentiation-related genes, such as ALP, osteocalcin (OCN), osterix and RUNX2, meanwhile, it inhibited the formation of the NLRP3 inflammasome.Citation124 Likewise, another compound caffeic acid phenethyl ester (CAPE) markedly suppressed osteogenic medium (OM)-induced calcification and decreased osteogenic gene/protein expression of RUNX2 and ALP in aortic VICs through the inhibition of NLRP3 inflammasome activation.Citation125
Clinical Medication
Amitriptyline, a synthetic tricyclic compound used as an antidepressant, has been found to reduce the expression of NLRP3 in OBs. This down-regulation of NLRP3 inhibited innate immune responses mediated by TLR4 and IL-1 receptors. As a result, amitriptyline showed potential for repurposing in the treatment of joint inflammatory conditions such as OA, both locally and systemically.Citation126 Similarly, by utilizing data text mining and computational pharmacology, Franco-Trepat et al discovered that naloxone and thalidomide were involved in downregulating the NLRP3 inflammasome pathway for the treatment of OA. These compounds have demonstrated anti-inflammatory and anti-catabolic properties in joint primary OA cells, including OBs.Citation127 As mentioned above, estrogen deficiency can induce NLRP3 inflammasome activation and impair OBs differentiation. A recent study demonstrated melatonin was found to enhance OBs differentiation in primary bone marrow MSCs obtained from mice with OVX. This effect was achieved through regulating Wnt/β-catenin signaling and the inhibition of NLRP3 inflammasome activation in femoral bone proteins and induced OBs stimulated by OVX.Citation18 Traditional Chinese medicine (TCM) is also playing a similar role in exerting its effects, Anemarrhenae Rhizoma/Phellodendri Chinensis Cortex (AR/PCC) herb pair has been extensively employed in TCM. Importantly, it was able to alleviate the abnormal activation of OBs pyroptosis in the vertebral bodies of diabetic rats by reducing the elevated expressions of NLRP3, ASC, caspase-1, GSDMD, and IL-1β through the enhancement of the antioxidant response protein Nrf2, while simultaneously reducing Keap1.Citation128
Inhibitors
The pyroptosis inhibitor known as NSA has been found to have a significant impact on the secretion of IL-6, TNF-α and IL-1β. In addition, it has been shown to reverse the effects of ATP/LPS on the activity of ALP and the mRNA expression of differentiation-related genes in OBs. However, when NLRP3 was overexpressed, these positive effects of NSA on OBs were abolished.Citation84 Ac-YVAD-cmk, another inhibitor of pyroptosis, effectively hindered the activation of NLRP3 triggered by DICER1 dysregulation and successfully restored the viability of OBs.Citation129 Similarly, osteogenic gene expression (RUNX2, Bglap, and Col1a) was significantly improved in the diabetes-induced skeletal loss after Ac-YVAD-cmk administration.Citation19 NF 39 was found to be a novel NLRP3 inhibitor, which could promote OBs differentiation via inhibiting NLRP3, thereby reducing the production of IL-1β dependent on NLRP3 in vitro.Citation130 Besides, classic NLRP3 inflammasome inhibitors such as CY-09, MCC950 have been demonstrated in multiple studies to alleviate the impact of NLRP3 inflammasome on OBs function and differentiation in diverse diseases.Citation20,Citation67,Citation72,Citation119 ALP inhibitor decreased ALP activity and mRNA expressions of BMP, RUNX2, OCN, OPN, phosphorylated ERK, IκBα, AKT, TNF-α and NLRP3, leading to an inhibition of the osteogenic differentiation of VICs.Citation131
Conclusion and Perspectives
The NLRP3 inflammasome is a sophisticated protein complex located in the cytoplasm, which forms upon cellular disruption. Its assembly triggers the activation of caspase-1, which facilitates the maturation and release of inflammatory cytokines, such as IL-1β and IL-18, as well as inflammatory cell death termed pyroptosis.Citation134,Citation135 It is important to note that dysregulated activation of NLRP3 can contribute to the development of chronic inflammation and have a significant impact on the progression of inflammatory diseases such as vascular disease,Citation136 diabetic cardiomyopathyCitation137 and autoimmune diseases.Citation138 OBs, which are responsible for constructing our skeletal framework, are remarkably adaptable and crucial cells that require precise regulation throughout their differentiation stages to ensure the appropriate development and balance of the skeletal system.Citation59,Citation139 An increasing body of evidence shows elevated levels of NLRP3 inflammasome in inflammatory conditions cause OBs pyroptosis and impair osteogenic differentiation, the release of numerous inflammatory cytokines and signaling molecules disrupt the balance of bone, affects the function of distant organs, and has the potential to exacerbate both local and systemic inflammation. Hence, targeting the NLRP3 inflammasome has the potential to be a future therapeutic approach for addressing the abnormal function and differentiation of OBs induced by pro-inflammatory cytokines (). This not only applies to bone metabolic disorders but also extends to other diseases that are influenced by abnormal cytokines involved in OBs differentiation, such as RUNX2, ALP, OCN, and osterix.
It should be borne in mind that the role of NLRP3 inflammasome-mediated pyroptosis in OBs and their differentiation varies in different diseases. In bone metabolic diseases related to inflammation, inhibiting NLRP3 inflammasome often promotes OBs differentiation, increases bone remodeling, and contributes to better bone development and growth. However, in vascular calcification and chronic kidney disease, NLRP3 inflammasome plays a beneficial role by reducing the expression of osteogenic markers, thus decreasing cell calcification including VICs, and alleviating the condition of related diseases. In addition, NLRP3 inflammasome can be activated by various stimuli, including oxidative stress, endoplasmic reticulum stress, pathogens, and epigenetic factors such as non-coding RNAs. Therefore, we speculate that targeting these initiating factors may lead to the development of new therapies for abnormal osteogenic differentiation and function caused by NLRP3 inflammasome activation. It is gratifying that research has been conducted on the development and screening of drugs targeting the upstream of NLRP3 inflammasome to treat related diseases.Citation140 Unfortunately, there is currently a lack of preclinical evidence demonstrating the correlation between NLRP3 inflammasome and OBs as well as osteogenic differentiation. Nevertheless, research both in vitro and in vivo has further bolstered our confidence in exploring NLRP3 inflammasome as a novel therapeutic approach for OBs-related diseases.
Abbreviations
OBs, osteoblasts; MSCs, mesenchymal stem cells; OC, osteoclasts; OP, osteoporosis; OA, osteoarthritis; OPN, osteopontin; RNUX2, runt-related transcription factor 2; PCD, programmed mode of cell death; IL, interleukin; PRRs, pattern recognition receptors; PAMPs, pathogen-associated molecular patterns; ASC, apoptosis-associated speckle-like protein; GSDMD, Gasdermin D; TLRs, toll-like receptors; LPS, lipopolysaccharide; NF-κB, nuclear factor-kappa B; PTM, post-translational modification; ALP, alkaline phosphatase; ROS, reactive oxygen species; OVX, ovariectomy; ATRA, all-trans retinoic acid; lncRMA, long non-coding RNA; miRNA, microRNA; VICs, valvular interstitial cells; ELP2, elongator complex protein 2; ART, artesunate; NAC, N-acetylcysteine; NSA, necrosulfonamide; BPA, bisphenol A; OCN, osteocalcin; CAPE, caffeic acid phenethyl ester; TCM, traditional Chinese medicine; AP/PCC, Rhizoma/Phellodendri Chinensis Cortex.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflicts of interest in this work.
Additional information
Funding
References
- Ponzetti M, Rucci N. Osteoblast differentiation and signaling: established concepts and emerging topics. Int J Mol Sci. 2021;22(13):6651. doi:10.3390/ijms22136651
- Chen W, Wu P, Yu F, Luo G, Qing L, Tang J. HIF-1α regulates bone homeostasis and angiogenesis, participating in the occurrence of bone metabolic diseases. Cells. 2022;11(22):3552. doi:10.3390/cells11223552
- Almalki SG, Agrawal DK. Key transcription factors in the differentiation of mesenchymal stem cells. Differentiation. 2016;92(1–2):41–51. doi:10.1016/j.diff.2016.02.005
- Eastell R, Szulc P. Use of bone turnover markers in postmenopausal osteoporosis. Lancet Diabetes Endocrinol. 2017;5(11):908–923. doi:10.1016/S2213-8587(17)30184-5
- Liu J, Zhang Y, Wu Y, et al. Delivery of m7G methylated Runx2 mRNA by bone-targeted lipid nanoparticle promotes osteoblastic bone formation in senile osteoporosis. Nano Today. 2024;54:102074. doi:10.1016/j.nantod.2023.102074
- Luo X, Wan Q, Cheng L, Xu R. Mechanisms of bone remodeling and therapeutic strategies in chronic apical periodontitis. Front Cell Infect Microbiol. 2022;12:908859. doi:10.3389/fcimb.2022.908859
- Zhu S, Zhu J, Zhen G, et al. Subchondral bone osteoclasts induce sensory innervation and osteoarthritis pain. J Clin Invest. 2019;129(3):1076–1093. doi:10.1172/JCI121561
- Tamaddon M, Gilja H, Wang L, et al. Osteochondral scaffolds for early treatment of cartilage defects in osteoarthritic joints: from bench to clinic. Biomate Rransl. 2020;1(1):3–17. doi:10.3877/cma.j.issn.2096-112X.2020.01.002
- Zheng G, Zhao Y, Li Z, et al. GLSP and GLSP-derived triterpenes attenuate atherosclerosis and aortic calcification by stimulating ABCA1/G1-mediated macrophage cholesterol efflux and inactivating RUNX2-mediated VSMC osteogenesis. Theranostics. 2023;13(4):1325–1341. doi:10.7150/thno.80250
- Singh P, Harris PC, Sas DJ, Lieske JC. The genetics of kidney stone disease and nephrocalcinosis. Nat Rev Nephrol. 2022;18(4):224–240. doi:10.1038/s41581-021-00513-4
- Pillai ICL, Li S, Romay M, et al. Cardiac fibroblasts adopt osteogenic fates and can be targeted to attenuate pathological heart calcification. Cell Stem Cell. 2017;20(2):218–232.e215. doi:10.1016/j.stem.2016.10.005
- Yang F, Liu S, Gu Y, et al. MicroRNA-22 promoted osteogenic differentiation of valvular interstitial cells by inhibiting CAB39 expression during aortic valve calcification. Cell Mol Life Sci. 2022;79(3):146. doi:10.1007/s00018-022-04177-6
- Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Sig Transd Target Therapy. 2021;6(1):128. doi:10.1038/s41392-021-00507-5
- Bergsbaken T, Fink SL, Cookson BT. Pyroptosis: host cell death and inflammation. Nat Rev Microbiol. 2009;7(2):99–109. doi:10.1038/nrmicro2070
- Swanson KV, Deng M, Ting JP. The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nat Rev Immunol. 2019;19(8):477–489. doi:10.1038/s41577-019-0165-0
- Wang Z, Zhou F, Feng X, et al. FoxO1/NLRP3 inflammasome promotes age-related alveolar bone resorption. J Dent Res. 2023;102(8):919–928. doi:10.1177/00220345231164104
- Detzen L, Cheat B, Besbes A, et al. NLRP3 is involved in long bone edification and the maturation of osteogenic cells. J Cell Physiol. 2021;236(6):4455–4469. doi:10.1002/jcp.30162
- Xu L, Zhang L, Wang Z, et al. Melatonin suppresses estrogen deficiency-induced osteoporosis and promotes osteoblastogenesis by inactivating the NLRP3 inflammasome. Calcified Tissue Int. 2018;103(4):400–410. doi:10.1007/s00223-018-0428-y
- Behera J, Ison J, Voor MJ, Tyagi N. Exercise-linked skeletal irisin ameliorates diabetes-associated osteoporosis by inhibiting the oxidative damage-dependent miR-150-FNDC5/Pyroptosis axis. Diabetes. 2022;71(12):2777–2792. doi:10.2337/db21-0573
- Lu J, Xie S, Deng Y, Xie X, Liu Y. Blocking the NLRP3 inflammasome reduces osteogenic calcification and M1 macrophage polarization in a mouse model of calcified aortic valve stenosis. Atherosclerosis. 2022;347:28–38. doi:10.1016/j.atherosclerosis.2022.03.005
- Pham TH, Kim EN, Trang NM, Jeong GS. Gallic acid induces osteoblast differentiation and alleviates inflammatory response through GPR35/GSK3β/β-catenin signaling pathway in human periodontal ligament cells. J Periodontal Res. 2023;59(1):204–219. doi:10.1111/jre.13208
- Martinon F, Burns K, Tschopp J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Molecular Cell. 2002;10(2):417–426. doi:10.1016/S1097-2765(02)00599-3
- Kayagaki N, Stowe IB, Lee BL, et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature. 2015;526(7575):666–671. doi:10.1038/nature15541
- Christgen S, Kanneganti TD. Inflammasomes and the fine line between defense and disease. Curr Opinion Immunol. 2020;62:39–44. doi:10.1016/j.coi.2019.11.007
- Bauernfried S, Hornung V. Human NLRP1: from the shadows to center stage. J Exp Med. 2022;219(1). doi:10.1084/jem.20211405
- Calabrese L, Fiocco Z, Mellett M, et al. Role of the NLRP1 inflammasome in skin cancer and inflammatory skin diseases. Br J Dermatol. 2024;190(3):305–315. doi:10.1093/bjd/ljad421
- Wen J, Xuan B, Liu Y, et al. Updating the NLRC4 inflammasome: from bacterial infections to autoimmunity and cancer. Front Immunol. 2021;12:702527. doi:10.3389/fimmu.2021.702527
- Duncan JA, Canna SW. The NLRC4 inflammasome. Immunol Rev. 2018;281(1):115–123. doi:10.1111/imr.12607
- Du L, Wang X, Chen S, Guo X. The AIM2 inflammasome: a novel biomarker and target in cardiovascular disease. Pharmacol Res. 2022;186:106533. doi:10.1016/j.phrs.2022.106533
- Man SM, Karki R, Malireddi RK, et al. The transcription factor IRF1 and guanylate-binding proteins target activation of the AIM2 inflammasome by Francisella infection. Nat Immunol. 2015;16(5):467–475. doi:10.1038/ni.3118
- Chae JJ, Wood G, Richard K, et al. The familial Mediterranean fever protein, pyrin, is cleaved by caspase-1 and activates NF-kappaB through its N-terminal fragment. Blood. 2008;112(5):1794–1803. doi:10.1182/blood-2008-01-134932
- Schnappauf O, Chae JJ, Kastner DL, Aksentijevich I. The pyrin inflammasome in health and disease. Front Immunol. 2019;10:1745. doi:10.3389/fimmu.2019.01745
- La Bella S, Di Ludovico A, Di Donato G, et al. The pyrin inflammasome, a leading actor in pediatric autoinflammatory diseases. Front Immunol. 2023;14:1341680. doi:10.3389/fimmu.2023.1341680
- Magnotti F, Chirita D, Dalmon S, et al. Steroid hormone catabolites activate the pyrin inflammasome through a non-canonical mechanism. Cell Rep. 2022;41(2):111472. doi:10.1016/j.celrep.2022.111472
- Lau R, Hann MM, Ottmann C. Crystal structure and ligandability of the 14-3-3/pyrin interface. Biochem Biophys Res Commun. 2023;651:1–7. doi:10.1016/j.bbrc.2023.02.013
- Mangan MSJ, Olhava EJ, Roush WR, Seidel HM, Glick GD, Latz E. Targeting the NLRP3 inflammasome in inflammatory diseases. Nat Rev Drug Discov. 2018;17(8):588–606. doi:10.1038/nrd.2018.97
- Latz E, Xiao TS, Stutz A. Activation and regulation of the inflammasomes. Nat Rev Immunol. 2013;13(6):397–411. doi:10.1038/nri3452
- Xia CY, Guo YX, Lian WW, et al. The NLRP3 inflammasome in depression: potential mechanisms and therapies. Pharmacol Res. 2023;187:106625. doi:10.1016/j.phrs.2022.106625
- Rashidi M, Wicks IP, Vince JE. Inflammasomes and cell death: common pathways in microparticle diseases. Trends Mol Med. 2020;26(11):1003–1020. doi:10.1016/j.molmed.2020.06.005
- Broz P, Dixit VM. Inflammasomes: mechanism of assembly, regulation and signalling. Nat Rev Immunol. 2016;16(7):407–420. doi:10.1038/nri.2016.58
- He Y, Hara H, Núñez G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem Sci. 2016;41(12):1012–1021. doi:10.1016/j.tibs.2016.09.002
- Zheng X, Wan J, Tan G. The mechanisms of NLRP3 inflammasome/pyroptosis activation and their role in diabetic retinopathy. Front Immunol. 2023;14:1151185. doi:10.3389/fimmu.2023.1151185
- Gritsenko A, Yu S, Martin-Sanchez F, et al. Priming is dispensable for NLRP3 inflammasome activation in human monocytes in vitro. Front Immunol. 2020;11:565924. doi:10.3389/fimmu.2020.565924
- Kodi T, Sankhe R, Gopinathan A, Nandakumar K, Kishore A. New insights on NLRP3 inflammasome: mechanisms of activation, inhibition, and epigenetic regulation. J Neuroimmune Pharmacol. 2024;19(1):7. doi:10.1007/s11481-024-10101-5
- Song N, Li T. Regulation of NLRP3 inflammasome by phosphorylation. Front Immunol. 2018;9:2305. doi:10.3389/fimmu.2018.02305
- Zhang Y, Luo L, Xu X, et al. Acetylation is required for full activation of the NLRP3 inflammasome. Nat Commun. 2023;14(1):8396. doi:10.1038/s41467-023-44203-0
- Xia J, Jiang S, Dong S, Liao Y, Zhou Y. The role of post-translational modifications in regulation of NLRP3 inflammasome activation. Int J Mol Sci. 2023;24(7):6126. doi:10.3390/ijms24076126
- Huang Y, Xu W, Zhou R. NLRP3 inflammasome activation and cell death. Cell Mol Immunol. 2021;18(9):2114–2127. doi:10.1038/s41423-021-00740-6
- Qiang R, Li Y, Dai X, Lv W. NLRP3 inflammasome in digestive diseases: from mechanism to therapy. Front Immunol. 2022;13:978190. doi:10.3389/fimmu.2022.978190
- Kelley N, Jeltema D, Duan Y, He Y. The NLRP3 inflammasome: an overview of mechanisms of activation and regulation. Int J Mol Sci. 2019;20(13):3328. doi:10.3390/ijms20133328
- Shi H, Wang Y, Li X, et al. NLRP3 activation and mitosis are mutually exclusive events coordinated by NEK7, a new inflammasome component. Nat Immunol. 2016;17(3):250–258. doi:10.1038/ni.3333
- Yue Z, Zhang X, Gu Y, et al. Regulation and functions of the NLRP3 inflammasome in RNA virus infection. Front Cell Infect Microbiol. 2023;13:1309128. doi:10.3389/fcimb.2023.1309128
- He XF, Li LL, Xian WB, et al. Chronic colitis exacerbates NLRP3-dependent neuroinflammation and cognitive impairment in middle-aged brain. J Neuroinflammation. 2021;18(1):153. doi:10.1186/s12974-021-02199-8
- Bai B, Yang Y, Wang Q, et al. NLRP3 inflammasome in endothelial dysfunction. Cell Death Dis. 2020;11(9):776. doi:10.1038/s41419-020-02985-x
- Chen Y, Yang Q, Lv C, et al. NLRP3 regulates alveolar bone loss in ligature-induced periodontitis by promoting osteoclastic differentiation. Cell Proliferation. 2021;54(2):e12973. doi:10.1111/cpr.12973
- Jiang N, An J, Yang K, et al. NLRP3 inflammasome: a new target for prevention and control of osteoporosis? Front Endocrinol. 2021;12:752546. doi:10.3389/fendo.2021.752546
- Karsenty G, Kronenberg HM, Settembre C. Genetic control of bone formation. Annu Rev Cell Dev Biol. 2009;25(1):629–648. doi:10.1146/annurev.cellbio.042308.113308
- Kim JM, Lin C, Stavre Z, Greenblatt MB, Shim JH. Osteoblast-osteoclast communication and bone homeostasis. Cells. 2020;9(9):2073. doi:10.3390/cells9092073
- Hu K, Olsen BR. Osteoblast-derived VEGF regulates osteoblast differentiation and bone formation during bone repair. J Clin Invest. 2016;126(2):509–526. doi:10.1172/JCI82585
- Wagley Y, Chesi A, Acevedo PK, et al. Canonical Notch signaling is required for bone morphogenetic protein-mediated human osteoblast differentiation. Stem Cells. 2020;38(10):1332–1347. doi:10.1002/stem.3245
- Harada S, Rodan GA. Control of osteoblast function and regulation of bone mass. Nature. 2003;423(6937):349–355. doi:10.1038/nature01660
- Komori T. Regulation of proliferation, differentiation and functions of osteoblasts by Runx2. Int J Mol Sci. 2019;20(7):1694. doi:10.3390/ijms20071694
- Liu SC, Sun QZ, Qiao XF, et al. LncRNA TUG1 influences osteoblast proliferation and differentiation through the Wnt/β-catenin signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(11):4584–4590. doi:10.26355/eurrev_201906_18035
- Shimada K, Crother TR, Karlin J, et al. Oxidized mitochondrial DNA activates the NLRP3 inflammasome during apoptosis. Immunity. 2012;36(3):401–414. doi:10.1016/j.immuni.2012.01.009
- Xian H, Watari K, Sanchez-Lopez E, et al. Oxidized DNA fragments exit mitochondria via mPTP- and VDAC-dependent channels to activate NLRP3 inflammasome and interferon signaling. Immunity. 2022;55(8):1370–1385.e1378. doi:10.1016/j.immuni.2022.06.007
- Sun D, Peng Y, Ge S, Fu Q. USP1 Inhibits NF-κB/NLRP3 induced pyroptosis through TRAF6 in osteoblastic MC3T3-E1 cells. J Musculoskelet Neuronal Interact. 2022;22(4):536–545.
- Liu S, Du J, Li D, et al. Oxidative stress induced pyroptosis leads to osteogenic dysfunction of MG63 cells. J Mol Histol. 2020;51(3):221–232. doi:10.1007/s10735-020-09874-9
- Lei L, Sun J, Han J, Jiang X, Wang Z, Chen L. Interleukin-17 induces pyroptosis in osteoblasts through the NLRP3 inflammasome pathway in vitro. Int Immunopharmacol. 2021;96:107781. doi:10.1016/j.intimp.2021.107781
- Khosla S, Oursler MJ, Monroe DG. Estrogen and the skeleton. Trends Endocrinol Metab. 2012;23(11):576–581. doi:10.1016/j.tem.2012.03.008
- Liang S, Nian Z, Shi K. Inhibition of RIPK1/RIPK3 ameliorates osteoclastogenesis through regulating NLRP3-dependent NF-κB and MAPKs signaling pathways. Biochem Biophys Res Commun. 2020;526(4):1028–1035. doi:10.1016/j.bbrc.2020.03.177
- Sasso G, Cerri PS, Sasso-Cerri E, Simões MJ, Gil CD, Florencio-Silva R. Possible role of annexin A1/FPR2 pathway in COX2/NLRP3 inflammasome regulation in alveolar bone cells of estrogen-deficient female rats with diabetes mellitus. J Periodontol. 2023. doi:10.1002/JPER.23-0530
- Guo L, Zhang Y, Liu H, Cheng Q, Yang S, Yang D. All-trans retinoic acid inhibits the osteogenesis of periodontal ligament stem cells by promoting IL-1β production via NF-κB signaling. Int Immunopharmacol. 2022;108:108757. doi:10.1016/j.intimp.2022.108757
- McCall SH, Sahraei M, Young AB, et al. Osteoblasts express NLRP3, a nucleotide-binding domain and leucine-rich repeat region containing receptor implicated in bacterially induced cell death. J Bone Miner Res. 2008;23(1):30–40. doi:10.1359/jbmr.071002
- Zhao P, Liu J, Pan C, Pan Y. NLRP3 inflammasome is required for apoptosis of Aggregatibacter actinomycetemcomitans-infected human osteoblastic MG63 cells. Acta Histochem. 2014;116(7):1119–1124. doi:10.1016/j.acthis.2014.05.008
- Ran S, Chu M, Gu S, Wang J, Liang J. Enterococcus faecalis induces apoptosis and pyroptosis of human osteoblastic MG63 cells via the NLRP3 inflammasome. Int Endodontic J. 2019;52(1):44–53. doi:10.1111/iej.12965
- Yoshida K, Okamura H, Hiroshima Y, et al. PKR induces the expression of NLRP3 by regulating the NF-κB pathway in Porphyromonas gingivalis-infected osteoblasts. Exp Cell Res. 2017;354(1):57–64. doi:10.1016/j.yexcr.2017.03.028
- Zhu X, Zhang K, Lu K, et al. Inhibition of pyroptosis attenuates Staphylococcus aureus-induced bone injury in traumatic osteomyelitis. Ann Translat Med. 2019;7(8):170. doi:10.21037/atm.2019.03.40
- Dai W, Wang M, Wang P, et al. lncRNA NEAT1 ameliorates LPS‑induced inflammation in MG63 cells by activating autophagy and suppressing the NLRP3 inflammasome. IntJ Mol Med. 2021;47(2):607–620. doi:10.3892/ijmm.2020.4827
- Yang Y, Miao L, Chang S, et al. Exosome-derived LncRNA TCONS_00072128 mediated osteogenic differentiation and inflammation by Caspase 8 regulation. Front Genetics. 2021;12:831420. doi:10.3389/fgene.2021.831420
- Yang Q, Zhao W, Chen Y, et al. RelA/MicroRNA-30a/NLRP3 signal axis is involved in rheumatoid arthritis via regulating NLRP3 inflammasome in macrophages. Cell Death Dis. 2021;12(11):1060. doi:10.1038/s41419-021-04349-5
- Crivaro A, Bondar C, Mucci JM, et al. Gaucher disease-associated alterations in mesenchymal stem cells reduce osteogenesis and favour adipogenesis processes with concomitant increased osteoclastogenesis. Mol Gene Metabol. 2020;130(4):274–282. doi:10.1016/j.ymgme.2020.06.003
- Xu L, Shen L, Yu X, Li P, Wang Q, Li C. Effects of irisin on osteoblast apoptosis and osteoporosis in postmenopausal osteoporosis rats through upregulating Nrf2 and inhibiting NLRP3 inflammasome. Exp Ther Med. 2020;19(2):1084–1090. doi:10.3892/etm.2019.8313
- Zhao W, Cao Y, Chen Y, et al. NLRP3 regulates mandibular healing through interaction with UCHL5 in MSCs. Int J Bio Sci. 2023;19(3):936–949. doi:10.7150/ijbs.78174
- Zhang J, Wei K. Necrosulfonamide reverses pyroptosis-induced inhibition of proliferation and differentiation of osteoblasts through the NLRP3/caspase-1/GSDMD pathway. Exp Cell Res. 2021;405(2):112648. doi:10.1016/j.yexcr.2021.112648
- Chen G, Deng C, Li YP. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int J Bio Sci. 2012;8(2):272–288. doi:10.7150/ijbs.2929
- Li K, Han J, Wang Z. Histone modifications centric-regulation in osteogenic differentiation. Cell Death Discovery. 2021;7(1):91. doi:10.1038/s41420-021-00472-6
- Guo J, Wang F, Hu Y, et al. Exosome-based bone-targeting drug delivery alleviates impaired osteoblastic bone formation and bone loss in inflammatory bowel diseases. Cell Rep Med. 2023;4(1):100881. doi:10.1016/j.xcrm.2022.100881
- Guo X, Wu Z. GABARAP ameliorates IL-1β-induced inflammatory responses and osteogenic differentiation in bone marrow-derived stromal cells by activating autophagy. Sci Rep. 2021;11(1):11561. doi:10.1038/s41598-021-90586-9
- Xie Z, Yu W, Zheng G, et al. TNF-α-mediated m(6)A modification of ELMO1 triggers directional migration of mesenchymal stem cell in ankylosing spondylitis. Nat Commun. 2021;12(1):5373. doi:10.1038/s41467-021-25710-4
- Bonar SL, Brydges SD, Mueller JL, et al. Constitutively activated NLRP3 inflammasome causes inflammation and abnormal skeletal development in mice. PLoS One. 2012;7(4):e35979. doi:10.1371/journal.pone.0035979
- Ahn JS, Seo Y, Oh SJ, et al. The activation of NLRP3 inflammasome potentiates the immunomodulatory abilities of mesenchymal stem cells in a murine colitis model. BMB Rep. 2020;53(6):329–334. doi:10.5483/BMBRep.2020.53.6.065
- Yang C, Song B, Han L, Gao Z. Study on the mechanism of NLRP3 effect on the skeleton of de-ovalized mice. Biochem Biophys Rep. 2023;35:101496. doi:10.1016/j.bbrep.2023.101496
- Ding Z, Chen W, Wu H, et al. Integrative network fusion-based multi-omics study for biomarker identification and patient classification of rheumatoid arthritis. Chin Med. 2023;18(1):48. doi:10.1186/s13020-023-00750-8
- Duan Y, Li H, Dong X, Geng Z, Xu X, Liu Y. VEGF mitigates bisphosphonate-induced apoptosis and differentiation inhibition of MC3T3-E1 cells. Exp Ther Med. 2022;23(2):130. doi:10.3892/etm.2021.11053
- Xia C, Ou S, Yang Y, et al. ELP2-NLRP3-GSDMD/GSDME-mediated pyroptosis is induced by TNF-α in MC3T3-E1 cells during osteogenic differentiation. J Cell Mol Med. 2023;27(24):4093–4106. doi:10.1111/jcmm.17994
- Fang M, Li B, Li X, Wang Y, Zhuang Y. MicroRNA-29b regulates pyroptosis involving calcific aortic valve disease through the STAT3/SOCS1 pathway. Int J Cardiol. 2023;371:319–328. doi:10.1016/j.ijcard.2022.08.029
- Jun C, Jun L, Shiting L, Qiangguo G, Gang Z. 降钙素基因相关肽通过抑制Nod样受体蛋白3表达促进小鼠成骨细胞分化的研究 [Calcitonin gene-related peptide inhibits the expression of Nod-like receptor protein 3 to Dromote osteoblast differentiation in mouse osteoblasts in vitro]. Hua xi kou qiang yi xue za zhi. 2016;34(1):12–16. Chinese. doi:10.7518/hxkq.2016.01.003
- Zheng W, Meng Z, Zhu Z, et al. Metal-organic framework-based nanomaterials for regulation of the osteogenic microenvironment. Small;2024. e2310622. doi:10.1002/smll.202310622
- Yang N, Liu Y. The role of the immune microenvironment in bone regeneration. Int J Med Sci. 2021;18(16):3697–3707. doi:10.7150/ijms.61080
- Zhang F, Lv M, Wang S, et al. Ultrasound-triggered biomimetic ultrashort peptide nanofiber hydrogels promote bone regeneration by modulating macrophage and the osteogenic immune microenvironment. Bioact Mater. 2024;31:231–246. doi:10.1016/j.bioactmat.2023.08.008
- Long J, Yao Z, Zhang W, et al. Regulation of osteoimmune microenvironment and osteogenesis by 3D-Printed PLAG/black phosphorus scaffolds for bone regeneration. Adv Sci. 2023;10(28):e2302539. doi:10.1002/advs.202302539
- Bai L, Song P, Su J. Bioactive elements manipulate bone regeneration. Biomate Rransl. 2023;4(4):248–269. doi:10.12336/biomatertransl.2023.04.005
- Zhang Y, Yan M, Niu W, et al. Tricalcium phosphate particles promote pyroptotic death of calvaria osteocytes through the ROS/NLRP3/Caspase-1 signaling axis in amouse osteolysis model. Int Immunopharmacol. 2022;107:108699. doi:10.1016/j.intimp.2022.108699
- Zhang Y, Yan M, Shan W, et al. Bisphenol A induces pyroptotic cell death via ROS/NLRP3/Caspase-1 pathway in osteocytes MLO-Y4. Food Chem Toxicol. 2022;159:112772. doi:10.1016/j.fct.2021.112772
- Yao Y, Cai X, Ren F, et al. The Macrophage-Osteoclast Axis in Osteoimmunity and Osteo-Related Diseases. Front Immunol. 2021;12:664871. doi:10.3389/fimmu.2021.664871
- Wang Z, Luo W, Zhang G, et al. FoxO1 knockdown inhibits RANKL-induced osteoclastogenesis by blocking NLRP3 inflammasome activation. Oral Dis. 2023. doi:10.1111/odi.14800
- An Y, Zhang H, Wang C, et al. Activation of ROS/MAPKs/NF-κB/NLRP3 and inhibition of efferocytosis in osteoclast-mediated diabetic osteoporosis. FASEB J. 2019;33(11):12515–12527. doi:10.1096/fj.201802805RR
- Yao L, Huang C, Dai J. Staphylococcus aureus enhances osteoclast differentiation and bone resorption by stimulating the NLRP3 inflammasome pathway. Mol Biol Rep. 2023;50(11):9395–9403. doi:10.1007/s11033-023-08900-9
- Alippe Y, Wang C, Ricci B, et al. Bone matrix components activate the NLRP3 inflammasome and promote osteoclast differentiation. Sci Rep. 2017;7(1):6630. doi:10.1038/s41598-017-07014-0
- Pieters BCH, Cappariello A, van den Bosch MHJ, van Lent P, Teti A, van de Loo FAJ. Macrophage-derived extracellular vesicles as carriers of alarmins and their potential involvement in bone homeostasis. Front Immunol. 2019;10:1901. doi:10.3389/fimmu.2019.01901
- Dai B, Xu J, Li X, et al. Macrophages in epididymal adipose tissue secrete osteopontin to regulate bone homeostasis. Nat Commun. 2022;13(1):427. doi:10.1038/s41467-021-27683-w
- Muñoz J, Akhavan NS, Mullins AP, Arjmandi BH. Macrophage Polarization and Osteoporosis: a Review. Nutrients. 2020;12(10):2999. doi:10.3390/nu12102999
- Zhu L, Wang Z, Sun X, et al. STAT3/Mitophagy axis coordinates macrophage NLRP3 inflammasome activation and inflammatory bone loss. J Bone Miner Res. 2023;38(2):335–353. doi:10.1002/jbmr.4756
- Chen Y, Wu Y, Guo L, et al. Exosomal Lnc NEAT1 from endothelial cells promote bone regeneration by regulating macrophage polarization via DDX3X/NLRP3 axis. J Nanobiotechnol. 2023;21(1):98. doi:10.1186/s12951-023-01855-w
- Hou L, Ye Y, Gou H, et al. A20 inhibits periodontal bone resorption and NLRP3-mediated M1 macrophage polarization. Exp Cell Res. 2022;418(1):113264. doi:10.1016/j.yexcr.2022.113264
- Ge G, Bai J, Wang Q, et al. Punicalagin ameliorates collagen-induced arthritis by downregulating M1 macrophage and pyroptosis via NF-κB signaling pathway. Sci China Life Sci. 2022;65(3):588–603. doi:10.1007/s11427-020-1939-1
- Yang L, Tao W, Xie C, et al. Interleukin-37 ameliorates periodontitis development by inhibiting NLRP3 inflammasome activation and modulating M1/M2 macrophage polarization. J Periodontal Res. 2024;59(1):128–139. doi:10.1111/jre.13196
- Grim JC, Aguado BA, Vogt BJ, et al. Secreted factors from proinflammatory macrophages promote an osteoblast-like phenotype in valvular interstitial cells. Arteriosclerosis Thrombosis Vasc Biol. 2020;40(11):e296–e308. doi:10.1161/ATVBAHA.120.315261
- Wang Z, Feng X, Zhang G, et al. Artesunate ameliorates ligature-induced periodontitis by attenuating NLRP3 inflammasome-mediated osteoclastogenesis and enhancing osteogenic differentiation. Int Immunopharmacol. 2023;123:110749. doi:10.1016/j.intimp.2023.110749
- Liu H, Zhang X, Zhong X, et al. Puerarin inhibits vascular calcification of uremic rats. Eur J Pharmacol. 2019;855:235–243. doi:10.1016/j.ejphar.2019.05.023
- Wang C, Xia Y, Qu L, Liu Y, Liu X, Xu K. Cardamonin inhibits osteogenic differentiation of human valve interstitial cells and ameliorates aortic valve calcification via interfering in the NF-κB/NLRP3 inflammasome pathway. Food Funct. 2021;12(23):11808–11818. doi:10.1039/D1FO00813G
- Franco-Trepat E, Alonso-Pérez A, Guillán-Fresco M, et al. β boswellic acid blocks articular innate immune responses: an in silico and in vitro approach to traditional medicine. Antioxidants. 2023;12(2):371. doi:10.3390/antiox12020371
- Kim EN, Nabende WY, Jeong H, Hahn D, Jeong GS. The marine-derived natural product epiloliolide isolated from sargassum horneri regulates NLRP3 via PKA/CREB, promoting proliferation and anti-inflammatory effects of human periodontal ligament cells. Mar Drugs. 2021;19(7):388. doi:10.3390/md19070388
- Chen Y, Hu W, Wang Y, et al. A selected small molecule prevents inflammatory osteolysis through restraining osteoclastogenesis by modulating PTEN activity. Clin transl med. 2020;10(8):e240. doi:10.1002/ctm2.240
- Liu M, Li F, Huang Y, et al. Caffeic acid phenethyl ester ameliorates calcification by inhibiting activation of the AKT/NF-κB/NLRP3 inflammasome pathway in human aortic valve interstitial cells. Front Pharmacol. 2020;11:826. doi:10.3389/fphar.2020.00826
- Franco-Trepat E, Alonso-Pérez A, Guillán-Fresco M, et al. Amitriptyline blocks innate immune responses mediated by toll-like receptor 4 and IL-1 receptor: preclinical and clinical evidence in osteoarthritis and gout. Br J Pharmacol. 2022;179(2):270–286. doi:10.1111/bph.15707
- Franco-Trepat E, Guillán-Fresco M, Alonso-Pérez A, et al. Repurposing drugs to inhibit innate immune responses associated with TLR4, IL1, and NLRP3 signaling in joint cells. Biomed Pharmacother. 2022;155:113671. doi:10.1016/j.biopha.2022.113671
- Fu F, Luo H, Du Y, et al. AR/PCC herb pair inhibits osteoblast pyroptosis to alleviate diabetes-related osteoporosis by activating Nrf2/Keap1 pathway. J Cell Mol Med. 2023;27(22):3601–3613. doi:10.1111/jcmm.17928
- Greene E, Flees J, Dhamad A, et al. Double-stranded RNA is a novel molecular target in osteomyelitis pathogenesis: a translational avian model for human bacterial chondronecrosis with osteomyelitis. Am J Pathol. 2019;189(10):2077–2089. doi:10.1016/j.ajpath.2019.06.013
- Chen W, Tang P, Fan S, Jiang X. A novel inhibitor INF 39 promotes osteogenesis via blocking the NLRP3/IL-1β axis. Biomed Res Int. 2022;2022:7250578. doi:10.1155/2022/7250578
- Zhang C, Liu M, Wang X, et al. ALP inhibitors inhibit inflammatory responses and osteoblast differentiation in hVIC via AKT-ERK pathways. Alternat Therap Health Med. 2023;29(1):58–65.
- Zeng XZ, Zhang YY, Yang Q, et al. Artesunate attenuates LPS-induced osteoclastogenesis by suppressing TLR4/TRAF6 and PLCγ1-Ca(2+)-NFATc1 signaling pathway. Acta Pharmacol Sin. 2020;41(2):229–236. doi:10.1038/s41401-019-0289-6
- Ye Y, Chen A, Li L, et al. Repression of the antiporter SLC7A11/glutathione/glutathione peroxidase 4 axis drives ferroptosis of vascular smooth muscle cells to facilitate vascular calcification. Kidney Int. 2022;102(6):1259–1275. doi:10.1016/j.kint.2022.07.034
- Wang L, Hauenstein AV. The NLRP3 inflammasome: mechanism of action, role in disease and therapies. Mol Aspect Med. 2020;76:100889. doi:10.1016/j.mam.2020.100889
- Elliott EI, Sutterwala FS. Initiation and perpetuation of NLRP3 inflammasome activation and assembly. Immunol Rev. 2015;265(1):35–52. doi:10.1111/imr.12286
- Takahashi M. NLRP3 inflammasome as a key driver of vascular disease. Cardiovasc Res. 2022;118(2):372–385. doi:10.1093/cvr/cvab010
- Yang F, Qin Y, Wang Y, et al. Metformin Inhibits the NLRP3 Inflammasome via AMPK/mTOR-dependent Effects in Diabetic Cardiomyopathy. Int J Bio Sci. 2019;15(5):1010–1019. doi:10.7150/ijbs.29680
- Zhang Y, Yang W, Li W, Zhao Y. NLRP3 inflammasome: checkpoint connecting innate and adaptive immunity in autoimmune diseases. Front Immunol. 2021;12:732933. doi:10.3389/fimmu.2021.732933
- Titorencu I, Pruna V, Jinga VV, Simionescu M. Osteoblast ontogeny and implications for bone pathology: an overview. Cell Tissue Res. 2014;355(1):23–33. doi:10.1007/s00441-013-1750-3
- Jin X, Liu D, Zhou X, Luo X, Huang Q, Huang Y. Entrectinib inhibits NLRP3 inflammasome and inflammatory diseases by directly targeting NEK7. Cell Rep Med. 2023;4(12):101310. doi:10.1016/j.xcrm.2023.101310