Abstract
Context: Ficus religiosa Linn (Moraceae) is a variety of fig tree. Its figs are known to contain a high serotonergic content, and modulation of serotonergic neurotransmission plays a crucial role in the pathogenesis of amnesia. Thus, the present study was envisaged.
Objective: To investigate the effect of the methanol extract of figs of Ficus religiosa (FRFE) on scopolamine-induced anterograde and retrograde amnesia in mice.
Materials and methods: Transfer latency (TL) to the preferred niche in the elevated plus-maze (EPM) and learning avoidance of passive behavior to avoid punishment in the modified passive avoidance paradigm (MPA) served as behavioral models for the assessment of memory. Scopolamine (1 mg/kg, i.p.) was administered before training for induction of anterograde amnesia and before retrieval for induction of retrograde amnesia in both models. TL in the EPM, step down latency (SDL), number of trials, and number of mistakes in the MPA were determined in vehicle control, FRFE treated (10, 50, and 100 mg/kg, i.p.), and standard groups (piracetam 200 mg/kg, i.p.). Cyproheptadine, a non-selective 5-HT1/2 blocker (4 mg/kg, i.p.), was administered along with the FRFE to investigate the involvement of serotonergic pathways in the anti-amnesic effect of FRFE.
Results and discussion: FRFE resulted in a significant improvement of memory, as its treatment attenuated the scopolamine-induced anterograde and retrograde amnesia dose-dependently. Further, cyproheptadine pretreatment significantly reversed the anti-amnesic effect of FRFE.
Conclusion: FRFE has anti-amnesic activity against scopolamine-induced amnesia, in a dose-dependent manner. Inhibition of the anti-amnesic effect of FRFE by cyproheptadine substantiates the involvement of serotonergic pathways for its activity.
Introduction
Amnesia refers to a specific, acquired difficulty in learning new information and/or retrieving information from the past. It is characterized by severe disruption of memory without deficits in intelligence, attention, perception, or judgments (CitationNelson et al., 1995; CitationMark et al., 2004). Memory is defined as a process of registration, storage, and retrieval of information, which occurs due to changes in transmission efficacy at the synapse where nerve cells communicate with each other (CitationKandel et al., 1991). There are two major classes of amnesia, anterograde and retrograde. Anterograde amnesia is impairment of memory for events occurring after the incident. In such a case, new memories are not formed. On the other hand, retrograde amnesia is impairment of memory of the events which have occurred before the incident; in such a case, new memories can be formed, but old memories are lost (CitationNelson et al., 1995; CitationMark et al., 2004). There are numerous factors responsible for the genesis of clinical amnesia; these include: increased oxidative stress (CitationGracia et al., 1997); alteration in brain neurotransmitter systems such as cholinergic (CitationCastellano et al., 1996), γ-aminobutyric acid (GABA)-ergic (CitationPitsikas et al., 2003), glutamatergic (CitationBrain et al., 2007), dopaminergic (CitationSawaguchi & Goldman, 1994), and nitrergic (CitationPrickaerts et al., 1997) systems. Apart from all these, the brain serotonergic system is known to play a vital role in learning and memory, as all serotonin receptors are densely expressed in brain regions associated with learning and memory and have been implicated in various human cognitive disorders directly or indirectly. Activation of these serotonin receptors results in modulation of brain cholinergic, glutamatergic, dopaminergic, GABAergic, and/or nitrergic systems (CitationBuhot, 1997; CitationBuhot et al., 2000; CitationTagliaferro et al., 2003; CitationMadeleine et al., 2008), which are directly involved in cognitive disorders. In general, decreased serotonergic neurotransmission in the brain is associated with severe memory impairment, and modulating its neurotransmission can ameliorate such memory impairment (Jolane & David, Citation2001).
Ficus religiosa Linn (Moraceae), commonly known as the Bodhi tree, is regarded as a sacred tree to both Hindus and Buddhists. It is recognized for its medicinal as well as religious purposes in India (CitationKala et al., 2006). Major active constituents present in its leaves include campestrol, stigmasterol, 28-isofucosterol, α-amyrin, β-amyrin, lupeol, tannic acid, n-nonacosane, hexacosanol, n-octacosanol, and amino acids such as l-cystine, l-lysine, l-arginine, dl-serine, dl-aspartic acid, glycine, dl-threonine, dl-alanine, l-proline, tryptophan, l-tyrosine, dl-methionine, dl-valine, and l-leucine (CitationVerma & Bhatia, 1986). The bark contains vitamin K1, n-octacosanol, methyl oleanolate, lanosterol, sitosterol, stigmasterol, bergapten, and bergaptol (CitationSwami et al., 1989; CitationSwami & Bisht, 1996). Figs (fruits) of Ficus religiosa contain numerous amino acids such as asparagine, tyrosine, alanine, threonine, and valine (CitationAli & Qadry, 1987). Apart from amino acids, figs of this tree have been reported to contain the highest amount of serotonin (5-HT), as compared to figs of other Ficus species (CitationBliebtrau, 1968). Moreover, the figs were also characterized in our previous study for the presence of serotonin (CitationSingh & Goel, 2009), as modulation of serotonergic neurotransmission in the brain plays a crucial role in higher cognitive processes. This led us to envisage that figs of Ficus religiosa may possess anti-amnesic activity via modulating brain serotonin levels, which would be of clinical usefulness.
Materials and methods
Preparation and characterization of extract
The extract was prepared and characterized as reported in our previous study (CitationSingh & Goel, 2009). Briefly, the figs were collected, washed with water, shade-dried, powdered, and subjected to extraction by refluxing with 70% methanol (1:10 w/v) in a Soxhlet extractor for 10–12 h. The resultant extract was evaporated to dryness using a rotary evaporator (Heidolph Laborota 4001), to obtain a dried Ficus religiosa fig extract (FRFE). Further, high performance liquid chromatography (HPLC) fingerprinting was employed to confirm the presence of serotonin in FRFE. The weighed amount of FRFE was reconstituted by dissolving it in dimethylsulfoxide (DMSO) and then suspending the resultant solution in 0.5% carboxymethyl cellulose (CMC) suspension (DMSO:CMC, 1:9) freshly before use, and was injected intraperitoneally (i.p.). Control groups received an equal volume of vehicle (DMSO:CMC, 1:9) i.p.
Drugs and chemicals
All standard chemicals used in this study were of analytical grade. Piracetam was obtained from UCB (Belgium), dimethylsulfoxide (DMSO) from Qualigens Fine Chemicals (Mumbai, India), carboxymethyl cellulose (CMC) and methanol from S.D.Fine-Chem Ltd. (Mumbai, India), cyproheptadine hydrochloride from Q.P. Pharma Chem (Dera Bassi, India), scopolamine and serotonin from Acrose (Belgium), and HPLC grade solvents from Spectrochem Pvt. Ltd. (India).
Animals
Swiss Albino mice (20–30 g) of either sex were employed in the present study, in different groups (n = 6). The animals were housed in standard cages and maintained at room temperature with natural day and night cycles. The animals were allowed free access to food (standard laboratory rodent’s chow) and water during the study period. The experiments were conducted between 7.00 and 16.00 h. The animals were acclimatized to the laboratory conditions 5 days prior to the behavioral study. All procedures were conducted according to guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals, India (CitationCPCSEA, 2003) and were approved by the Institutional Ethical Committee.
Elevated plus-maze paradigm
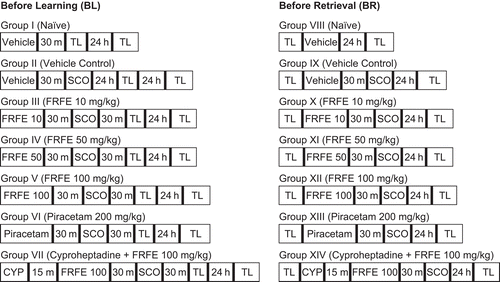
The elevated plus-maze served as an exteroceptive behavioral model to evaluate learning and memory in mice (CitationItoh et al., 1991). The elevated plus-maze for mice consisted of two open arms (16 cm × 5 cm) and two closed arms (16 cm × 5 cm × 12 cm) and the maze was elevated to a height of 25 cm from the floor. The animals were divided into 14 groups (Groups I–XIV) and were treated as shown in . On the first day, each mouse was placed at the end of an open arm, facing away from the central platform, and transfer latency (TL) was recorded, which is defined as time (seconds) taken by the animal to move from an open arm into one of the closed arms with all four legs. The mouse was allowed to explore the maze for another 2 min and then returned to the home cage. Retention of this learned task was examined 24 h after the first-day trial. Scopolamine (1 mg/kg, i.p.) was administered before training for induction of anterograde amnesia and before retrieval for induction of retrograde amnesia, and TL was recorded. A significant decrease in TL compared to the vehicle control group was considered as an anti-amnesic effect.
Figure 1. Treatment schedule and division of groups for elevated plus-maze paradigm. m, minutes; TL, transfer latency; h, hours; SCO, scopolamine (1 mg/kg, i.p.); FRFE 10, 50, and 100, Ficus religiosa fig extract (10, 50, and 100 mg/kg, i.p.); CYP, cyproheptadine (4 mg/kg, i.p.).
Modified passive avoidance paradigm
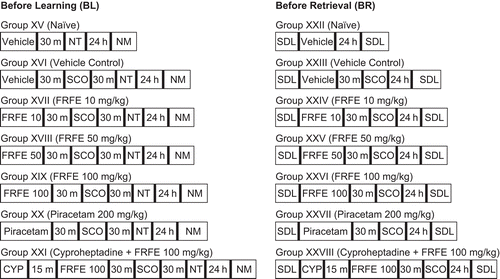
Passive avoidance behavior based on negative reinforcement was employed to examine long-term memory (CitationJarvik & Kopp, 1967). The apparatus consisted of a Plexiglas box (27 cm × 27 cm × 27 cm) with a grid floor (3 mm stainless steel rods set 8 mm apart), having a shock free zone (SFZ) (wooden platform 10 cm × 7 cm × 1.7 cm) in the center of the grid floor. Electric shock (20 V AC) was delivered to the grid floor. The animals were divided into 14 groups (Groups XV–XXVIII) and were treated as shown in . Each mouse was trained to stay on the SFZ for at least 90 s; for this the animals were gently placed on the wooden platform, and when the mouse stepped down placing all paws on the grid floor, shocks were delivered for 15 s and the step down latency (SDL) was recorded. SDL is defined as the time (seconds) taken by the mouse to step down from the wooden platform to the grid floor with all paws. The process was repeated several times until the animal learned to stay on the SFZ for at least 90 s. The number of trials required to learn this task was noted. Retention was tested after 24 h, whereby each mouse was again placed on the SFZ, and the SDL and numbers of mistakes were recorded, with an upper cut-off time of 120 s, for 5 min. Scopolamine (1 mg/kg, i.p.) was administered before training for the induction of anterograde amnesia and before retrieval for inducing retrograde amnesia. A significantly decreased number of mistakes and trials and increased SDL compared to the vehicle control group were considered as anti-amnesic effects.
Figure 2. Treatment schedule and division of groups for modified passive avoidance paradigm. m, minutes; SDL, step down latency; NT, number of trials; NM, number of mistakes; h, hours; SCO, scopolamine (1 mg/kg, i.p.); FRFE 10, 50, and 100, Ficus religiosa fig extract (10, 50, and 100 mg/kg, i.p.); CYP, cyproheptadine (4 mg/kg, i.p.).
Statistical analysis
Results are expressed as mean ± standard error of the mean (SEM). The data were subjected to one-way analysis of variance (ANOVA) followed by Newman–Keuls multiple comparison test. Results were regarded as significant at p < 0.05.
Results
Effect of FRFE on scopolamine-induced amnesia in elevated plus-maze
Effect on anterograde amnesia
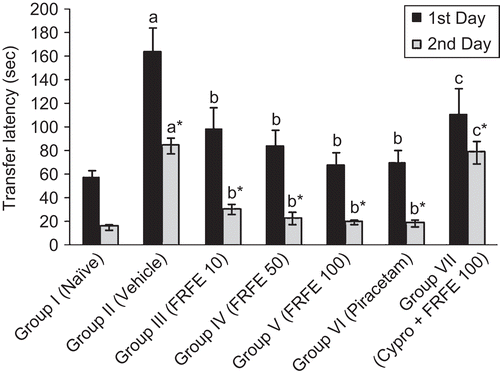
Scopolamine (1 mg/kg, i.p.) administered before learning significantly (p < 0.05) increased the TL in Group II (vehicle control) as compared to Group I (naive), on both the first and second days, indicating the induction of anterograde amnesia. On the other hand, the animals pretreated with FRFE (Groups III, IV, and V) showed a dose-dependent reduction in TL on both days compared to Group II (vehicle control), indicating a significant (p < 0.05) attenuation of anterograde amnesia. Piracetam at a dose of 200 mg/kg (Group VI) significantly (p < 0.05) improved memory and reversed the anterograde amnesia induced by scopolamine as compared to Group II (vehicle control). FRFE (100 mg/kg) treatment along with cyproheptadine (Group VII) failed to show an anti-amnesic effect, as this group showed a significant (p < 0.05) increase in TL as compared to Group V (FRFE 100 mg/kg per se treated group) ().
Figure 3. Effect of FRFE on transfer latency in scopolamine-induced amnesia (before learning). a,a*p < 0.05 as compared to Group I (naive) 1st and 2nd day respectively; b,b*p < 0.05 as compared to Group II (vehicle control) 1st and 2nd day respectively, and c,c*p < 0.05 as compared to Group V (FRFE 100 mg/kg) 1st and 2nd day respectively.
Effect on retrograde amnesia
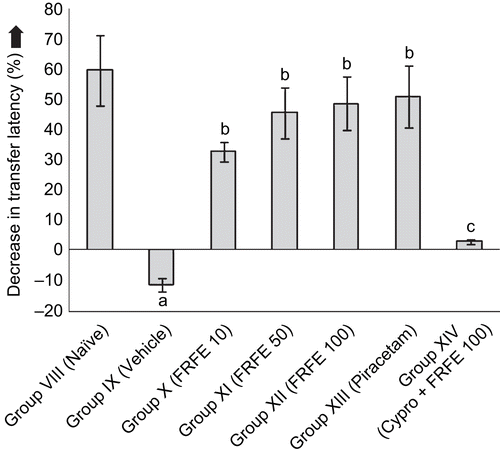
Scopolamine administered before retrieval showed the induction of retrograde amnesia, as Group IX (vehicle control) exhibited a significant (p < 0.05) increased TL on the second day as compared to Group VIII (naive), as there was a negative change in transfer latency. FRFE treatment (Groups X, XI, and XII) significantly (p < 0.05) attenuated increased TL, in a dose-dependent manner, when compared to Group IX (vehicle control). Cyproheptadine significantly (p < 0.05) attenuated the anti-amnesic effect of FRFE, as the combined treated group (Group XIV) showed a significant increase in TL as compared to the FRFE (100 mg/kg) per se treated group (Group XII) ().
Figure 4. Effect of FRFE on transfer latency in scopolamine-induced amnesia (before retrieval). ap < 0.05 as compared to Group VIII (naive), bp < 0.05 as compared to Group IX (vehicle control), and cp < 0.05 as compared to Group XII (FRFE 100 mg/kg).
Effect of FRFE on scopolamine-induced amnesia in modified passive avoidance paradigm
Effect on anterograde amnesia
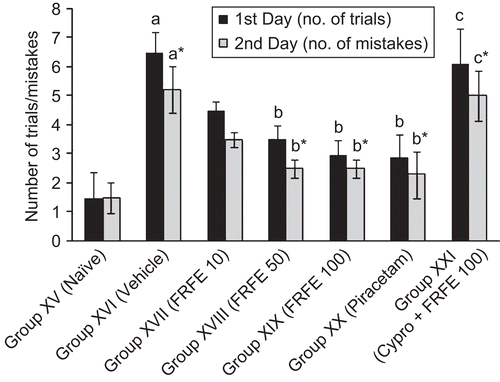
Scopolamine administered before learning significantly (p < 0.05) increased the number of trials on the first day and number of mistakes on the second day in Group XVI (vehicle control) as compared to Group XV (naive), indicating the induction of anterograde amnesia. Piracetam (Group XX) and FRFE (Groups XVIII and XIX) treatment significantly (p < 0.05) decreased the number of trials on the first day and mistakes on the second day as compared to Group XVI (vehicle control). However, the extract at a dose of 10 mg/kg (Group XVII) failed to show a significant decrease in trials and mistakes as compared to Group XVI (vehicle control). The anti-amnesic effect of FRFE was antagonized by pretreatment with cyproheptadine (Group XXI), as this group showed a significant (p < 0.05) increase in the number of trials on the first day and number of mistakes on the second day, as compared to the FRFE (100 mg/kg) per se treated group (Group XIX) ().
Figure 5. Effect of FRFE on number of trials and mistakes in scopolamine-induced amnesia (before learning). a,a*p < 0.05 as compared to Group XV (naive) 1st and 2nd day respectively, b,b*p < 0.05 as compared to Group XVI (vehicle control) 1st and 2nd day respectively, and c,c*p < 0.05 as compared to Group XIX (FRFE 100 mg/kg) 1st and 2nd day respectively.
Effect on retrograde amnesia
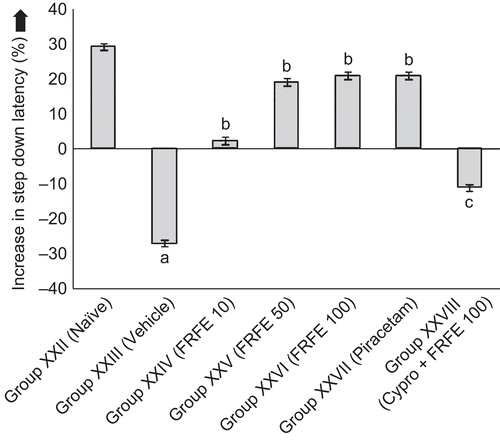
Scopolamine administered before retrieval significantly (p < 0.05) decreased the SDL in Group XXII (vehicle control) on the second day, as compared to Group XXIII (naive), as there was a negative change in SDL, showing induction of retrograde amnesia. FRFE treatment (Groups XXIV, XXV, and XXVI) significantly (p < 0.05) increased SDL, in a dose-dependent manner, when compared to Group XXIII (vehicle control). Cyproheptadine significantly (p < 0.05) attenuated the anti-amnesic effect of FRFE (100 mg/kg) (Group XXVIII), as compared to the FRFE (100 mg/kg) per se treated group (Group XXVI) ().
Figure 6. Effect of FRFE on step down latency in scopolamine-induced amnesia (before retrieval). ap < 0.05 as compared to Group XXII (naive), bp < 0.05 as compared to Group XXIII (vehicle control), and cp < 0.05 as compared to Group XXVI (FRFE 100 mg/kg).
Discussion
The role of brain neurochemical systems in cognitive functioning is a subject of increasing research interest, as cognitive disorders emerge as major public health problems (CitationWilliam & Thomas, 1991). The treatment strategies for amnesia involve reduction of oxidative stress (CitationGracia et al., 1997), enhancement of cholinergic tone (CitationBartus et al., 1982), increased release of nitric oxide (CitationPitsikas et al., 2002), decreased GABAergic activity (CitationMcNamara & Skelton, 1996), increased glutamatergic tone (CitationHlinak & Krejci, 2003), and increased dopaminergic (CitationHotte et al., 2005) and serotonergic functions (CitationSteckler & Sahgal, 1995). The literature has shown that brain serotonergic modulation affects all the abovementioned pathways directly or indirectly (CitationBuhot, 1997; CitationBuhot et al., 2000; CitationTagliaferro et al., 2003; CitationJaun et al., 2006; CitationMadeleine et al., 2008). It was hypothesized that memory impairment can be treated and/or prevented via targeting the brain serotonergic pathway. Based upon this, the present study was designed owing to the high serotonergic content of figs of Ficus religiosa. First, bias reports claiming the presence of the serotonergic component in figs were confirmed using the HPLC fingerprinting method (CitationSingh & Goel, 2009).
The results revealed in our study were concordant with previous findings (CitationStephan et al., 1995) that scopolamine induces anterograde and retrograde amnesia, as scopolamine increased the TL in EPM when injected before training and after training; it increased the number of trials on the first day and mistakes on the second day when injected before training, and decreased the SDL when injected after training in the MPA paradigm, as compared to the respective naive groups. FRFE treatment significantly attenuated the scopolamine-induced anterograde and retrograde amnesia in a dose-dependent manner and showed maximum anti-amnesic activity at 100 mg/kg; FRFE-treated groups showed a decrease in TL in the EPM when injected before training and after training. Similarly, FRFE treatment decreased the number of trials on the first day and mistakes on the second day when injected before training, and increased the SDL when injected after training in the MPA paradigm, when compared with the respective vehicle control groups. The anti-amnesic effect of FRFE at a dose of 100 mg/kg was comparable to that of piracetam (200 mg/kg). Cyproheptadine (non-selective 5-HT1/2 antagonist) attenuated the anti-amnesic effect of FRFE. This attenuation indicated the involvement of the serotonergic system in the anti-amnesic mechanism of action of FRFE.
From the results of the present study it is concluded that FRFE treatment attenuated scopolamine-induced anterograde and retrograde amnesia in a dose-dependent manner. Antagonism of the anti-amnesic effect of FRFE with cyproheptadine pretreatment showed that FRFE might be mediating its effect via modulating the serotonin-dependent activation of muscarinic cholinergic receptors (AChRs) (CitationStone et al., 1988; CitationBuhot, 1997; CitationBuhot et al., 2000). Further modulation of central cholinergic, nitrergic, and oxidative status by FRFE is under investigation to reveal the precise downstream signaling.
Declaration of interest: The authors report no conflict of interests. The authors alone are responsible for the content and writing of this paper.
References
- Ali M, Qadry JS (1987): Amino acid composition of fruits and seeds of medicinal plants. J Indian Chem Soc 64: 230–231.
- Bartus RT, Dean RL, Beer B, Lippa AS (1982): The cholinergic hypothesis of geriatric memory dysfunction. Science 217: 408–414.
- Bliebtrau JN (1968): The Parable of the Beast. New York, Macmillan Company, pp. 74.
- Brain MB, James NR, Glen TP, Robert MD, Robert JS, Gregory RJT (2007): Cognitive deficits in rats after fore brain cholinergic depletion are reversed by novel no mimetic nitrate ester. Neuropsychopharmacology 32: 505–513.
- Buhot MC, Martin S, Segu L (2000): Role of serotonin in memory impairment. Ann Med 32: 210–221.
- Buhot MC (1997): Serotonin receptors in cognitive behaviors. Curr Opin Neurobiol 7: 243–254.
- Castellano C, Cabid S, Puglisi AS (1996): Psychopharmacology of memory modulation: evidence for multiple interactions among neurotransmitters and hormones. Behav Brain Res 77: 1–21.
- CPCSEA (2003): CPCSEA guidelines for laboratory animal facility. Indian J Pharmacol 35: 257–274.
- Gracia R, Musleh W, Tocco G, Thompson RF, Baudry M (1997): Time dependent blockade of STP and LTP in hippocampal slices following acute stress in mice. Neurosci Lett 233: 41–44.
- Hlinak Z, Krejci I (2003): n-Methyl-d-aspartate prevented memory deficits induced by MK-801 in mice. Physiol Res 52: 809–812.
- Hotte M, Naudon L, Jay TM (2005): Modulation of recognition and temporal order memory retrieval by dopamine D1 receptor in rats. Neurobiol Learn Mem 84: 85–92.
- Itoh J, Nabeshima T, Kameyama T (1991): Utility of an elevated plus-maze for dissociation of amnesic and behavioral effects of drugs in mice. Eur J Pharmacol 194: 71–76.
- Jarvik ME, Kopp R (1967): An improved one-trial passive avoidance learning situation. Psychol Rep 21: 221–224.
- Jaun R, Pedro M, Francisco JP, Inmaculada B, Maria CM, Jordi M, Isaac T (2006): Role of serotonin in cerebral oxidative stress in rats. Acta Neurobiol Exp 66: 1–6.
- Jolane AB, David NF (2001): Tryptophan depletion and its implications for psychiatry. Br J Psychiatry 178: 399–405.
- Kala CP, Dhyani PP, Sajwan BS (2006): Developing the medicinal plants sector in northern India: challenges and opportunities. J Ethnobiol Ethnomed 2: 32–46.
- Kandel ER, Schwartz JH, Jessel TM (1991). Principles of Neural Science. New York, Elsevier, pp. 1009–1031.
- Madeleine VK, Charles AM, Kevin CFF (2008): A role for the 5-HT1A, 5-HT4 and 5-HT6 receptors in learning and memory. Trends Pharmacol Sci 29: 482–492.
- Mark K, Judy C, Larry RS (2004): Functional amnesia: clinical description and neuropsychological profile of 10 cases. Learn Mem 11: 213–226.
- McNamara KR, Skelton RW (1996): Baclofen, a selective GABAB receptor agonist, dose-dependently impairs spatial learning in rats. Pharmacol Biochem Behav 53: 303–308.
- Nelson B, Dean CD, John AL (1995): Clinical assessment of memory disorders in amnesia and dementia. Annu Rev Psychol 46: 493–523.
- Pitsikas N, Rigamonti AE, Cella SG, Muller EE (2002): Effects of the nitric oxide donor molsidomine on different memory components as assessed in the object-recognition task in the rat. Psychopharmacology 162: 239–245.
- Pitsikas N, Rigamonti AE, Cella SG, Muller EE (2003): The GABAB receptor and recognition memory: possible modulation of its behavioral effects by nitrergic system. Neuroscience 118: 1121–1127.
- Prickaerts J, Steinbush HW, Smits JF, DeVent J (1997): Possible role of nitric oxide cyclic GMP pathway in objection recognition memory: effects of 7-nitroindazole and zaprinast. Eur J Pharmacol 337: 125–136.
- Sawaguchi T, Goldman RPS (1994): The role of D1-dopamine receptor in working memory: local injections of dopamine antagonists into the prefrontal cortex of rhesus monkeys performing an oculomotor delayed-response task. J Neurophysiol 71: 515–528.
- Singh D, Goel RK (2009): Anticonvulsant effect of Ficus religiosa: role of serotonergic pathways. J Ethnopharmacol 123: 330–334.
- Steckler T, Sahgal A (1995): The role of serotonergic-cholinergic interactions in the mediation of cognitive behavior. Behav Brain Res 67: 165–199.
- Stephan GA, Stephen M, Michael SF (1995): Scopolamine selectively disrupts the acquisition of contextual fear conditioning in rats. Neurobiol Learn Mem 64: 191–194.
- Stone WS, Croul CE, Gold PE (1988): Attenuation of scopolamine-induced amnesia in mice. Psychopharmacology 96: 417–420.
- Swami KD, Bisht NPS (1996): Constituents of Ficus religiosa and Ficus infectoria and their biological activity. J Indian Chem Soc 73: 631.
- Swami KD, Malik GS, Bisht NPS (1989): Chemical investigation of stem bark of Ficus religiosa and Prosopis spicigera. J Indian Chem Soc 66: 288–289.
- Tagliaferro P, Ramos AJ, Lopez-Costa JJ, Lopez EM, Brusco A (2003): Changes in the postnatal development on nitric oxide system induced by serotonin depletion. Brain Res Dev Brain Res 146: 39–49.
- Verma RS, Bhatia KS (1986): Chromatographic study of amino acids of the leaf protein concentrates of Ficus religiosa Linn and Mimusops elengi Linn. Indian J Hosp Pharm 23: 231–232.
- William JM, Thomas HC (1991): Serotonin, memory, and the aging brain. Psychopharmacology 103: 143–149